- Information by Map Unit
- Information by Topic
- Documents in SoLo
Links to other related sites
- About SoLo
-
- Mission
- Disclaimer
- Authors (complete list)
Presettlement fire played an important role in nutrient conversion, plant succession, diversity, and stand dynamics in coniferous forests of western North America. Prescribed fire can maintain site quality and contribute to control of insect and disease problems while reducing wildfire hazard. Fire effects on soils are largely governed by interactions between fuel consumption and soil characteristics that influence soil heating. Many impacts on vegetation and site productivity are also related to soil heating.
Before the coming of western civilization, fire played an important role in nutrient dynamics, in plant succession, and in shaping stand density of coniferous forests in western North America (Arno 1980). Lightning was a common source of ignition, and Native Americans commonly used fire to manipulate grasslands and forests (Barrett and Arno 1982; Gruell 1985). However, Europeans who settled the West viewed fire as a destructive agent (Pyne 1982). The fires of 1910 in the Northern Rocky Mountain region greatly influenced Forest Service fire policy (Pyne 1982), and resulted in an obsession to suppress all fires. Beneficial uses of fire were largely ignored until the 1970's (Pyne 1982).
Concepts of multiple use and sustained yield are traditions in American forestry; but concerns about wildlife, threatened and endangered species, global change, and forest health have led to new demands on forests. The "new perspectives in forestry" concept raises additional questions about long-term productivity of forests. The total amount of plant material produced by a forest per unit area per year (net primary productivity) is a measure of forest productivity that integrates environmental site conditions, vegetation, and stage of vegetation development (Grier and others 1989). A better understanding of site productivity is needed to assess the effects of management activities, including fire's effect on factors that influence productivity.
It is clear that soil is a major determinant of site productivity and environmental site conditions are both extrinsic and intrinsic (Grier and others 1989). Extrinsic site factors (climate, topography, and geology) determine long-term regional productivity potentials and set the upper limit of site productivity. Intrinsic factors such as soil moisture-holding capacity, soil nutrient status, and soil porosity affect plant growth and productivity potential. Natural disturbances such as fire, insects, disease, and erosion alter intrinsic factors, influencing soil forming processes, vegetation development, and site productivity. Potentially, vegetation development and site productivity can be influenced by the use of fire, timber harvesting, and chemical application.
The purpose of this paper is to review the interactions between fire, vegetation, and soils that influence site productivity. We will first examine historic fire occurrence: frequency, size, and characteristics. Next, we will discuss variations in heat output from fire and how it affects soil heating and in turn, intrinsic site factors. Finally, we will discuss how fire affects the physical, chemical, and biological systems of the soil.
Study of historic fire frequencies, sizes, and effects provides insights into the ecological role of fire and how it influences forest productivity. Since fire is a natural part of the development of coniferous forests in western North America, the productive nature of the ecosystems is likely adapted to fire. There is considerable variation in relationships between fire occurrence, site productivity, and fuel accumulation. Decomposition and nutrient cycling processes in some ecosystems were controlled by recurring fire. In other ecosystems fire was relatively infrequent and herbivory and fungi assumed greater roles in decomposition and nutrient cycling (Gosz 1981; Olson 1981; Woodmansee and Wallach 1981). At the landscape level, fire potential can be generalized on the basis of productivity and physiognomy (fig. 1). Sites that are severely moisture or temperature limited are unproductive. These sites were fuel limited, and burned infrequently in the past (Martin 1982; Ryan in press). Such sites were dominated by herbaceous plants or shrubs, but few trees. Higher temperature and moisture in mesic coniferous forests increased productivity and fire frequency. In highly productive, but more moist ecosystems, large amounts of burnable biomass accumulated, but meteorological conditions were seldom favorable for extensive burning.
Figure 1—General relationship between site productivity, fuel loadings, and fire cycles in the intermountain West. Site productivity versus habitat type trends are excerpted from Pfister and others (1977). Fuel loadings versus productivity trends are adapted from Brown and See (1981). Fire cycles were adapted from Martin (1982). [view larger image - 20K] [Text description of this figure]
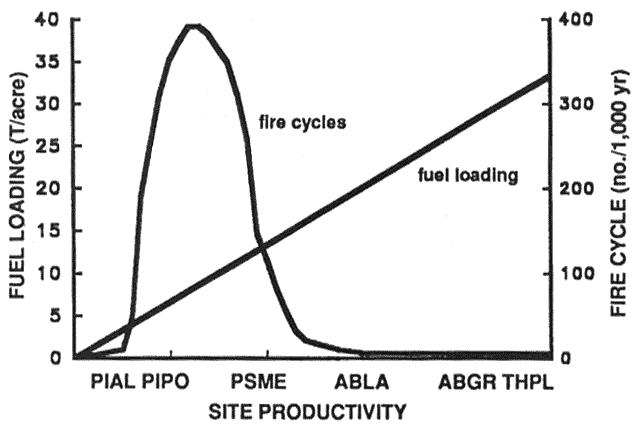
Biomass increases with time until it is removed by fire, logging, or grazing, or until litter accumulation and decomposition are in equilibrium (Gosz 1981; Olson 1981). As fire frequency increases, aboveground biomass approaches some lower limit capable of sustaining a fire (Martin 1982; Olson 1981). In western coniferous forests, moisture deficits vary from chronic on arid and less productive sites to infrequent on the most mesic sites. However, even on the most mesic sites, moisture deficits were frequent enough that fires likely consumed live and dead biomass before litter accumulation and decomposition were in equilibrium.
In low-elevation forests, light surface fires routinely spread from rangelands and seral grasslands. Fires burned through cured herbaceous fuels as often as every other year (Martin 1982), but fire frequencies of 5 to 20 years were more typical (Arno 1980; Martin 1982; Steele and others 1986). Fires often top killed shrubs and reduced encroachment of young conifers into rangelands (Gruell 1986; Gruell and others 1986), but they rarely killed the scattered large conifers. Such fires produced minimal soil heating (Ryan and Noste 1985). Frequent fires maintained open "parklike" stands of ponderosa pine (Pinus ponderosa) with well-developed herbaceous understories. Allocation of site water and nutrients among a few widely spaced trees resulted in vigorous growth and, apparently, relatively high resistance to insects and disease (Christiansen and others 1987; Crookston and Stark 1985).
During the early settlement of the West, extensive grazing removed much of the herbaceous fine fuel (Gruell 1986) in lower elevation forests. These forests were easily accessible and extensively logged. Logging and grazing altered the role of fire as an ecosystem process. In the absence of periodic surface fire these forests gradually accumulated fuels (Brown 1985; Ryan and Frandsen in press) and became overstocked with numerous dense thickets of young trees and shrubs (Arno 1988; Gruell 1986; Gruell and others 1982). These forests frequently experienced severe insect and disease problems that caused mortality and created fuels.
In the lower montane zone, fires burned on the average of every 20 to 50 years (Arno 1980; Martin 1982). At higher elevations fire frequencies of 50 to 100 years were more common. Surface fires commonly burned beneath stands, injuring and killing some trees and igniting smoldering ground fires (Ryan and Frandsen in press). Soil heating was variable as a result of spatial variation in ground and surface fuels. These surface fires released nutrients and reduced competition from late-successional and climax species. Stand replacing crown fires occurred during dry years where fuels had accumulated, resulting in patches of non forest or young forests across the landscape.
In the subalpine forests, presettlement fires were less frequent than at lower elevations. On productive sites, the fire regime intergraded with that of the montane forests. On less productive, drier, or colder sites, the minimum time to accumulate enough dead biomass to carry a fire in the absence of high wind was 20 to 50 years. Fires generally burned surface herbaceous fuels, frequently torching clumps of trees. Even light surface fires resulted in high mortality of the typically fire sensitive subalpine conifers (Ryan and Reinhardt 1988). Fire suppression in subalpine forests may not have had a major effect on fuels, species composition, or stand structure; but it probably affected the age distribution of subalpine forests.
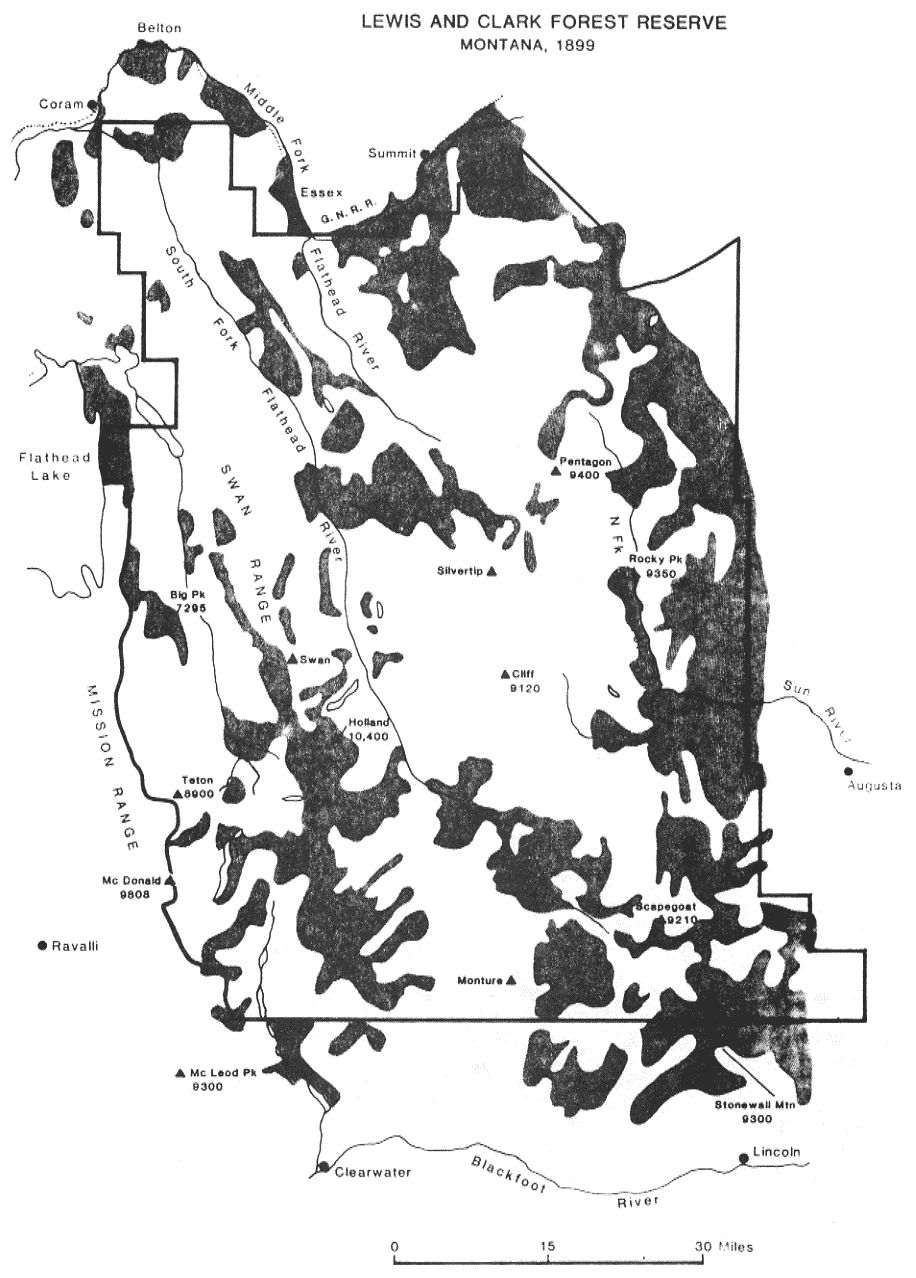
The prevalence of fire on the landscape was repeatedly observed by 19th century explorers. Evidence of frequent large fires was observed by Ayres (1901) in the Lewis and Clark Forest Reserve of Montana (includes the Bob Marshall and Scapegoat Wilderness complex) and by Leiberg (1899) in the Bitterroot Forest Reserve. Nearly a million acres (31 percent of the area) burned in the Lewis and Clark Reserve (fig. 2) between 1859 and 1898 (Ayres 1901). The 1889 fire in the Lewis and Clark Reserve burned about 800,000 acres in a stand-replacement fire. Leiberg estimated that 40 percent of the Bitterroot Reserve burned during the same period. Barrett (1982) found that some areas of Cook Mountain, in the Clearwater National Forest in Idaho, burned as many as five times between 1869 and 1919.
Some areas of the 1889 fire reburned in 1895, 1910, and again in the 1988 Canyon Creek fire (250,000 acres). Fires similar in size to the Canyon Creek fire historically occurred on the average of every 25 years in this area (Losensky 1990).
Figure 2—Areas burned in the 40-year period from 1860 to 1899 in the Lewis and Clark Forest Reserve. The shaded areas represent stand-replacement fires. Areas that underburned are not indicated. Map is redrawn from Ayres (1901). [view larger image - 108K] [Text description of this figure]
Many of the large fires of the 19th and early 20th century consumed all the forest floor, leaving the soil surface bare. These observations suggest that, historically, the burn interval was short enough to preclude the accumulation of forest floor depths typical of what we see today. Large areas of the forest were apparently maintained in earlier stages of succession than what we typically see today (Gruell 1983; Losensky 1990) (fig. 3). The effects of fire on site productivity remain uncertain, but it is apparent that fire played a role in nutrient cycling, species composition, and stocking control in virtually all western coniferous forests.
Figure 3—This photo comparison contrasts the appearance of a forest in 1900 (A) and in 1981 (B). The photos were taken from a ridge about 5 miles west of Haystack Butte, looking southwest across Smith Creek toward Crown Mountain on the east front of the Rocky Mountains, Lewis and Clark National Forest. In (A) the near slopes are in early succession following wildfire in the late 1800's that removed conifers and stimulated shrubs. In (B) the near slope in 1981 is now densely covered by Douglas-fir. Photos are taken from Gruell (1983): (A) is USGS photograph 665 by C. D. Walcott, and (B) is a photograph by G. E. Gruell. [view larger image of A - 84K] [view larger image of B - 76K] [Text description of A] [Text description of B]
A.

B.
Fire characteristics vary with fuels, weather, and topography. Some fires exhibit flaming, others smolder. Flames associated with forest fires are the result of the oxidation of pyrolysis products released from heated materials. Smoldering is pyrolysis and oxidation without flames. Crown fires, surface fires, and smoldering ground fires have different heat release and spread rate characteristics. Each fire type results in different levels of fuel consumption and above- and belowground heating. Surface fires are confined to litter, twigs, branches, and larger woody fuels on the forest floor. Live vegetation may add to the surface fire depending on its flammability. Fine surface fuels make up a small portion of the fuel loading on most sites, but their ignition and consumption determine fire spread, intensity, and ignition of other fuels. Rate of fire spread is primarily dependent on the moisture content of the fine fuels and wind speed. A light surface fire passing through needles, twigs, and small branches will not significantly heat either the crown foliage or mineral soil but will dry surface fuels. Surface fuels, twigs, cones, and logs act as ignition sources for the forest floor. Larger fuels are of less importance to fire spread, but contribute heat for the initiation of crown fires, drying of fuels, and the downward flow of heat into the mineral soil.
Flame length is a visual measure (Byram 1959) of fire intensity, which is the rate of energy released by the fire. If fire intensity is sufficiently high, flames may reach into the canopy and ignite the tree crown. Less intense surface fires may ignite understory trees and dead branches in the lower crown and carry the flames into the crown. Crowning and torching result in rapid spread rates and intense heat release for short durations, which results in little soil heating. Smoldering fires have low intensity and occur in tightly packed fuels of low moisture content. The tightly packed fuels on the forest floor are the fermentation (F) and humus (H) layers, which are made up of partly to well-decomposed organic matter. The F and H layers are roughly equivalent to the Oe and Oa horizons defined by the Soil Conservation Service, U.S. Department of Agriculture, (Federer 1982). In this paper, we will refer to the F and H layers combined as duff. The litter (L) layer is not part of the duff. Smoldering occurs in the forest floor duff (F and H layers) where packing ratios (volume fraction of fuel) are greater than 10 percent (Frandsen 1989). In contrast, surface litter packing ratios are less than 10 percent and exhibit flaming.
The term fire severity has been used to describe fire effects based on the appearance of the litter, duff, and soil after burning (Wells and others 1979). Severity reflects the degree of organic matter consumption and soil heating (Morgan and Neuenschwander 1988; Ryan and Noste 1985), but is ambiguous in quantification. In many instances, the term fire severity has been used synonymously with fire intensity. Present usage is confusing. In this paper we will discuss soil heating and the relationships to soil chemical, physical, and biological properties.
Heat flux describes the rate of heat flow that is delivered to the plant or the soil horizon of interest. The duration of heat flow and the resultant temperatures directly affect the soil-vegetation system (Wells and others 1979). Since we know much about the chemical, physical, and biological responses to specific temperatures, it is useful to express heat flow as temperature profiles when explaining fire effects (Harvey and others 1989).
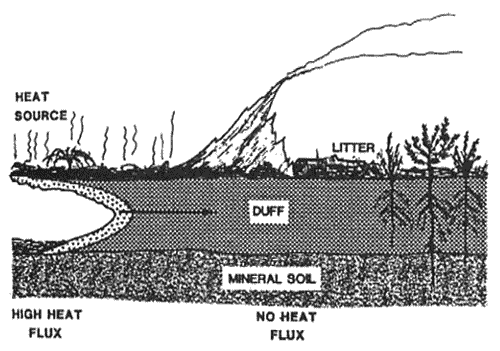
Rate of burning and heat output for surface and crown fires are influenced by fuel load, fuel moisture, topography, ignition method, air temperature, wind, and relative humidity. Heat output and burn rate for smoldering are influenced, in part, by the fuel moisture and inorganic content of the duff (Frandsen 1989). Rates of spread for flaming are a thousand times greater than for smoldering. When duff moisture content is high, heat from the surface fire may dry, ignite, and burn a portion of the duff (Ottmar and others 1985; Sandberg 1980), but only minimal soil heating results. When dry duff is present, smoldering is the primary source of heat (except for large fuel concentrations) to the lower duff and mineral soil (fig. 4). Although temperatures (500 to 600° C) from smoldering duff are lower than for flaming (1000 to 1500° C), the long duration of smoldering and the close proximity of duff to the soil results in greater heating. When duff does not burn, it becomes a barrier to heat flow to the mineral soil (Frandsen and Ryan 1986).
Figure 4—Diagram showing heat flux to the mineral soil where duff burning is the heat source. When duff does not burn, it is a barrier to heat flow. The arrow represents the direction of spread for duff burning at a rate of about 3 cm/hour. [Text description of this figure]
Attempts to understand soil heating by characterizing fire intensity have not been successful. Fire treatments have been characterized according to fire intensity (reaction intensity and fire line intensity) (Chandler and others 1983), visual estimates (Ryan and Noste 1985; Tarrant 1956a), heat fluxes, and amount and rate of fuel consumption (Albini 1975). A major problem with predicting soil heating is the inability to quantitatively relate prefire conditions and fire characteristics to the downward heat pulse. Rowe (1983) noted that release of energy as the fire passes may bear little relation to degree of soil heating as indicated by survival of plant parts buried in the soil. Shearer (1975) also showed that fire intensity alone was not closely related to soil heating as indicated by root mortality. Moisture content of the duff and mineral soil and duration of the fire were also important. Ryan and Noste (1985) stated that factors other than fire intensity determine the downward heat pulse. Models such as the duff consumption model (Van Wagner 1972) and the duff burnout model (Albini 1975) have not been useful for predicting slash burn heat release (Wells and others 1979). The formulation of a purely physical model describing downward heat flux to the litter and soil would be valuable (Wells and others 1979).
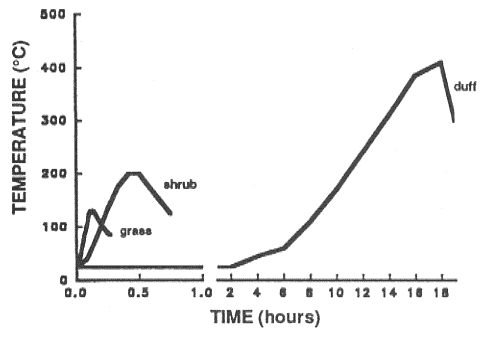
Maximum soil temperatures and duration of heating (fig. 5) varied between grass, shrub, and forest fires where duff burned (DeBano and others 1979; Frandsen 1989; Scotter 1970; Wells and others 1979). Soil heating and temperatures also varied within fires (DeBano and others 1979; Shearer 1975; Wells and others 1979). DeBano and others (1979) constructed stylized curves to predict soil temperatures for light, moderate, and intense chaparral fires. Models based on heat-transfer theory (Bristow and others 1986; DeVries 1958; Wierenga and DeWitt 1970) were developed to predict heat flow. Scotter (1970) attempted to model heat transfer and predict soil temperatures beneath fires, but did not include moisture-aided heat transfer. Aston and Gill (1976), Campbell and others (1990), Chinanzvavana and others (1986), and Pafford and others (1985) modeled the downward heat pulse in soil beneath fires. Chinanzvavana and others (1986), Pafford (1986), and Pafford and others (1985) evaluated heat-transfer processes between the fire and the underlying soil surfaces. These studies provide useful insights into the process of soil heating, but the models remain untested and cannot be used to predict the effect of fire on site productivity.
Figure 5—Maximum temperatures in the upper 2 cm of the mineral soil under a grass fire (Scotter 1970), a shrub fire (DeBano and others 1979), and a duff fire (Frandsen 1990). [Text description of this figure]
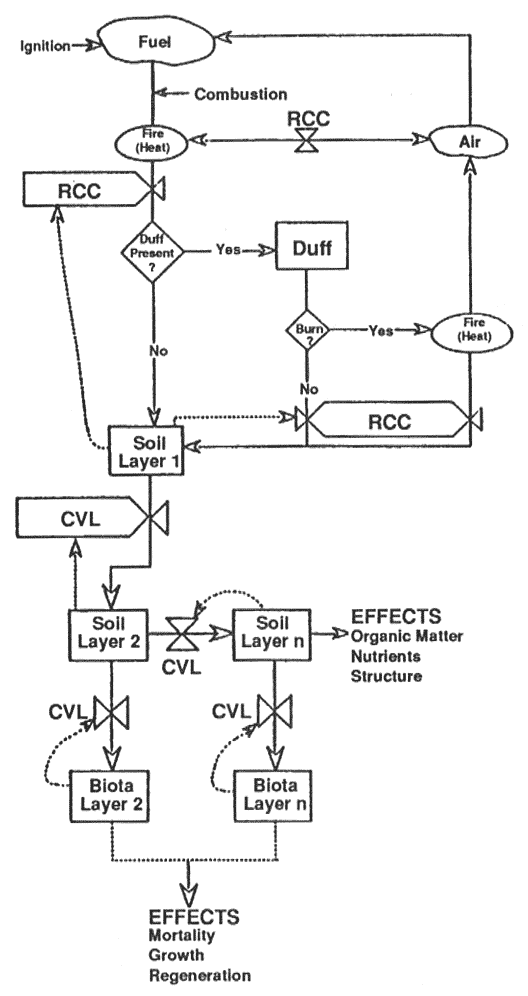
Heat flow from a fire through the soil-biota system is illustrated (fig. 6) by a conceptual model (Hungerford 1990). Heat generated from surface or ground fires is lost upward, transferred downward to the soil, or transferred downward to the duff if duff is not involved in the burn. Models of heat flow across the boundary layer between a fire and the soil are complex because of abrupt and dynamic changes in factors affecting heat transfer. Heat from the fire is transferred to unburned fuels, the atmosphere (air in fig. 6), and the soil or litter surface by radiation, convection, and conduction. Convection and radiation dominate, while conduction is important within a fuel element and between elements in contact. Only 8 to 10 percent (maximum of 25 percent) of the heat from burning is transmitted downward in the soil or litter (DeBano and others 1977; Packham 1969; Raison and others 1986; Steward 1989). The remaining 75 plus percent is lost upward and affects aboveground biological systems.
Figure 6—Model diagram of downward heat pulse from fire into the soil and biological systems. Rectangular boxes represent variables and solid lines between the boxes represent heat flow between the boxes. The processes that control heat flow between the boxes are shown by the "bow ties" on the solid lines. The dotted lines to the "bow ties" represent factors of the variables that influence the processes and rates of heat transfer between the boxes. [view larger image - 28K] [Text description of this figure]
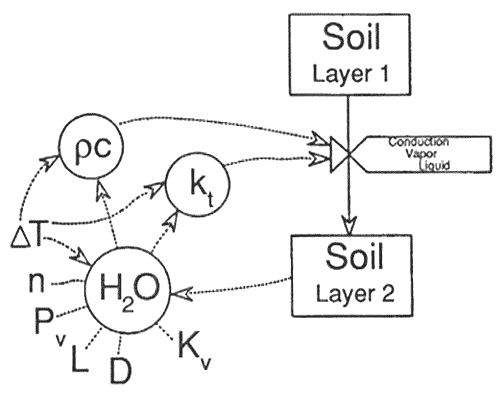
Heat transfer by conduction is important, but at soil temperature gradients created by fire, vapor flux accounts for the majority of heat transfer (Campbell and others 1990) and was as high as 60 percent (Westcot and Wierenga 1974). Liquid flux probably contributes little (10 percent or less) to heat transfer. Heat-transfer rates are influenced by numerous variables (fig. 7). Soil properties such as porosity, mineral content, water content, and organic content influence thermal conductivity, heat capacity, vapor conductivity, and hydraulic conductivity. It is clear from examination of the factors affecting heat transfer that knowledge of more than the fire characteristics is necessary for predicting soil heating and fire effects.
Figure 7—Details of the factors that control heat transfer between soil layers. The circles represent the primary variables (H2O = moisture content, kt = thermal conductivity, pc = heat capacity) that influence the processes of heat transfer (conduction, vapor movement, and liquid movement) between the layers. T = the temperature gradient, n = the vapor flow enhancement factor, Pv = the vapor density, L = the latent heat of vaporization, D = vapor diffusivity, and Kv = hydraulic conductivity; these are secondary variables that determine state of the primary variables. [Text description of this figure]
Little is known about heat transfer in living tissue. Most articles discuss heat transfer between the atmosphere and aboveground plants, particularly leaves (Campbell 1977; Gates 1980; Raschke 1960), or transfer within the stems of trees (Derby and Gates 1966; Herrington 1969; Martin 1963). Many of the variables that influence heat transfer in the soil (fig. 7) also influence heat transfer within living structures. The size (surface to volume ratio) and character of the biological structure affect heat capacity and thermal conductivity, thus influencing the final temperature. Thermal equilibrium with the soil likely will be established rather quickly for small structures, such as fungi and other microorganisms, and small plant parts. For larger structures such as roots, large rhizomes, tubers, and root crowns, temperatures may be different from the soil; thus heat transfer analysis is needed to evaluate the effects of soil heating.
Fire effects on the physical, chemical, and biological components of the duff and litter and mineral soil depend in part on the amount of fuel consumed. Fuel consumption depends on several factors, but moisture content has the strongest influence (Sandberg and Ottmar 1983). Other key factors that determine consumption in a specific fuel type are preburn amounts, mineral soil incorporation, degree of consumption of adjacent fuels, weather conditions, and ignition technique. Because the largest quantity of fuel (organic matter) at most northern Rocky Mountain sites occurs as duff and large woody material (Brown and See 1981), the influence of fire on these fuel groups is of great interest. These fuel types are potential sources of soil heating during burning and provide much of the nutrient base that determines long-term site productivity (Harvey and others 1989). Thus, controlling the degree of consumption by prescribed burning is important for controlling soil heating and potential site productivity.
In western larch/Douglas-fir (Larix occidentalis Nutt./Pseudotsuga menziesii var. glauca [Beissn.] Franco) clearcuts in northwestern Montana, Shearer (1975) found duff reduction by burning was closely related to moisture content. Sandberg (1980) used NFDRS 1,000-hour moisture content as a surrogate for duff moisture to predict duff consumption in Oregon and Washington Douglas-fir shelterwoods. Norum (1976) and Harrington (1987) reported duff consumption in western larch/Douglas-fir and ponderosa pine underburns, respectively, to be related to duff moisture and preburn duff amounts. Consumption of surface woody fuels was also important in Norum's (1976) duff-reduction model.
Large woody fuels are important for maintenance of long-term site productivity (Harvey and others 1987), but their consumption by fire is highly variable. Several factors influencing consumption of large woody fuels were identified, but their predictive value has not been developed. Moisture content was a key variable (Sandberg and Ottmar 1983). Degree of rot and grouping of fuel pieces are also influential factors (Albini 1976).
A wide range of fuel consumption from prescribed fires and wildfires can be expected because of the variability in factors that affect consumption. In a series of underburns in western larch/Douglas-fir in western Montana, large woody fuel reduction ranged from 0 to 41 tons/acre; small woody fuel reduction ranged from 0 to 6 tons/acre, and duff reduction ranged from 23 to 73 percent of the preburn depths, which averaged from 1.7 to 4.3 inches (Norum 1976). In several clearcut burns in northwestern Montana, duff depth reduction varied from 25 to 93 percent of the 2.5-inch preburn average (Shearer 1975). In northern Idaho, a series of slash underburns in ponderosa pine shelterwood cuts resulted in 0 to 17 tons/acre reduction of large woody fuels and 0 to 56 percent reduction of the duff, which averaged less than 3 inches before burning (Brown and others 1985). Different amounts of fuel consumption are expected to have different short- and long-term impacts on forested sites.
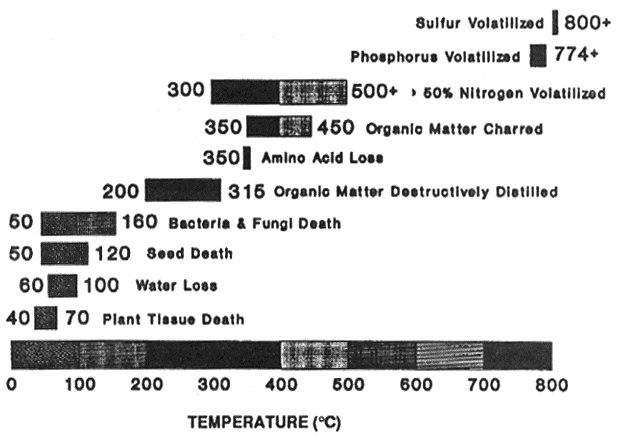
Combustion of organic matter is a rapid decomposition process in which oxidation changes the form of carbon (C), nitrogen (N), and other mineral elements (Harvey and others 1989). Most of the N found in forest systems is in an organic form that is unavailable for plant use. Physical and microbial decomposition are the general processes by which organic N is transformed into mineral N, but these processes are slow. The products of both biological and combustion oxidation can be similar. In both cases, the result is a mineralization of most nutrients. However, burning results in volatilization of N, phosphorus (P), and sulphur (S) (Wright and Bailey 1982). The degree of volatilization and subsequent loss of nutrients depend on the temperatures produced during burning (DeBano and others 1979) and the amount of duff consumed (Little and Ohman 1988) (fig. 8).
Figure 8—Temperature range for important temperature-sensitive events. [view larger image - 32K] [Text description of this figure]
Some of the N volatilized from the duff, however, is mineralized and condensed in the soil (Jurgensen and others 1981; Stark 1977). Direct losses of N from burning ponderosa pine ranged from 70 lb/acre in moderate ponderosa pine underburns (Covington and Sackett 1984) to 800 lb/acre in a severe wildfire (Grier 1975). Other elemental nutrients can be increased in the residual organic matter or soil (Viro 1974), can remain relatively unchanged (Nissley and others 1980), or can be reduced (Feller 1988) depending on the consumption and soil heating.
Fire effects on the physical and chemical properties of soil depend on the amount of material consumed during burning, the magnitude and duration of soil heating, the frequency of fire, and the postfire environment. High soil heating causes nutrient loss from the soil. Redistribution or losses of organics and nutrients within the soil often change the physical properties of the soil (Wells and others 1979).
Soil Structure and Water Absorption—In general, most fires do not cause enough soil heating to produce direct changes in soil structure. But if temperatures of 400 to 800° C occur the clay fraction in the soil is altered, making the texture more coarse and erodible (Chandler and others 1983). In some cases soil tilth was improved under the high temperatures of slash burning, because heating made clay more friable (Wells and others 1979). When duff is completely consumed, mineral soil is exposed to higher postfire temperatures and raindrop action, which often reduces soil porosity (Wells and others 1979). DeByle (1981) reported that burning increased bulk density of the soil on burns that varied widely in intensity and degree of soil heating.
The ability of soils to absorb water after fire is directly related to the degree of soil heating and amount of organic matter consumed. Infiltration rate is most often reduced by fire on forest and range soils, but some studies showed increased infiltration rates or no change (Wells and others 1979). Neal and others (1965) suggested that as soil heating increases, soil organic matter decreases, and moisture-holding capacity is reduced. A water-repellent layer is sometimes created when volatiles from burning organic materials are driven downward and condense on soil particles (DeBano and others 1979). Coarse-textured soils became more water repellent than fine-textured soils because the surface area of coarse grains is less than that for fine grains. Water movement and water holding capacity are reduced at and below the water-repellent layer. Formation of these water-repellent layers is an important concern on shrublands in California, Arizona, and Oregon (Chandler and others 1983), but is of less importance in the northern coniferous forests (DeByle 1981).
Soil Nutrients—Fire can alter the amount of organic matter and mineral nutrients in the soil just as it does in the duff and slash layers. Organic matter in the soil is destructively distilled between 200 and 300° C, is charred between 300 and 400° C, and is consumed above 450° C (fig. 8). Dyrness and Youngberg (1957) found a 60 percent reduction in organic matter in the upper 2 inches of soil on a portion of a clearcut burn in Oregon. DeByle (1976) reported a 12 percent loss of organics in the surface soils 1 year following several clearcut burns in western Montana, but preburn levels were reached by the second year. Within a clearcut burn in northern Idaho, Niehoff (1985) found the organic matter content in the surface 3 inches of soil to be only 0.6 percent on extremely burned microsites compared to 3.8 percent in unburned soils. Only 7 percent of the area, however, was classified as extremely burned.
A redistribution of organic matter from the forest floor or slash layers into the mineral soil may occur with light to moderate burning (Wells and others 1979). Increases in soil organic matter were reported for red pine (Pinus resinosa Ait.) underburning (Alban 1977) and Douglas-fir clearcut burning (Neal and others 1965). Other soil properties are affected by soil organic matter changes. As soil organic matter increases, total N increases along with other minerals (Lunt 1951; Wells and others 1979). Cation exchange capacity can also increase (Alban 1977). High soil heating may, however, reduce both of these properties (Wright and Bailey 1982). In light to moderate fuel-reduction burns, total N is frequently unchanged for ponderosa pine underburns (Kovacic and others 1986) and for Douglas-fir/western larch clearcut burns (Jurgensen and others 1981). Where soil temperatures reach 300° C and above (fig. 8), from 50 to 100 percent of the N will be volatilized (White and others 1973).
Cations are generally increased in the soil following their oxidation from burning the surface organic matter (Wells and others 1979). Potassium (K), calcium (Ca), magnesium (Mg), and phosphorus (P) were either increased or unchanged by underburns in ponderosa pine (Harrington 1977), red pine (Alban 1977), and Douglas-fir/western larch (Stark 1977). Losses of these minerals are possible only with high soil heating because volatilization temperatures are greater than 750° C (Wright and Bailey 1982). With the addition of cations (K, Ca, Mg), commonly acidic forest soils frequently experience an increase in pH (Wells and others 1979). This serves to further increase nutrient availability as well as improve the microenvironment for biological mineralization.
Nitrogen is a key element for plant growth and sites may experience reduced productivity when N is tied up in accumulated duff and fuels. Generally, fire causes an immediate increase in ammonium ions (NH4+), a readily available form of N (Jurgensen and others 1981). Ammonium levels were elevated for 6 months (Neal and others 1965) to 1 year after fire (Harrington 1977). With the addition of readily available nutrients and an increase in pH, as well as improved moisture and temperature conditions (Bisset and Parkinson 1980), biological mineralization and nitrification can increase on burned sites as long as carbon products are available (Wells and others 1979). These same microenvironment changes can also be favorable for symbiotic (Wright and Bailey 1982) and nonsymbiotic (Harvey and others 1979) N-fixation. Normally, the postfire flush of available nutrients returns to preburn levels with biological and physical immobilization and as the C:N ratio increases. Covington and Sackett (1986) reported that repeated interval underburning in ponderosa pine resulted in additional pulses of available N. If burns consume all of the forest floor and much of the soil organic matter, it follows that mineralization potential can be greatly reduced (Niehoff 1985). Because many stands are N limited, reduced mineralization can limit productivity (Harvey and others 1987).
Erosion—Forest organic matter, both living and dead, is important in controlling soil erosion. Excessive removal of vegetation and forest floor horizons increases erosion potential. Actual soil movement depends on storm intensity, slope steepness, and soil type (Wright and Bailey 1982), but the surface, nutrient-ladened soils are the first to be lost. Under extremely dry conditions where most of the forest floor and deep-rooted vegetation is removed by fire, erosion potential is greatly increased. Campbell and others (1977) estimated that a severe storm following a ponderosa pine wildfire removed 13.8 tons of soil per acre. Within 1 year, returning ground cover significantly reduced erosion. Following clearcutting and prescribed burning in a Douglas-fir/western larch cover type, erosion from snowmelt and summer storms was temporarily increased (Packer and Williams 1976). With the exception of the most intense burns on south slopes, soil movement was reduced within 3 years. Vegetation recovery and organic matter deposition on bare soils, are important for reducing erosion potential. Slower vegetation recovery is associated with greater soil heating (Lyon 1966; Morgan and Neuenschwander 1988), which affects the length of time the soil is susceptible to erosion.
Plant adaptations to climate and disturbance, including fire, were grouped and classified by several investigators (Grime 1979; Lyon and Stickney 1976; Noble and Slatyer 1977; Raunkiaer 1934; Rowe 1983). These classifications are working hypotheses of how species adaptations function, and were used as a basis for some plant succession models (Cattelino and others 1979; Hungerford 1987; Keane 1987; Morgan and Neuenschwander 1988) and for describing plant response to fire (Fischer 1989; Fischer and Bradley 1987). The relative resistance of plant species to damage from fire depends on the degree and duration of soil heating, the depth of perennating plant parts, heat resistance, and colonization potential (Morgan and Neuenschwander 1988; Noste and Bushey 1987). The following discussion reviews lethal temperature thresholds for plants, factors that influence damage, and species rooting habits.
Lethal Temperature and Heat Thresholds—The concept of stress resistance (Levitt 1980) is useful for understanding plant responses to heat. Stress resistance represents the ability of plants to survive and adjust to stress. A plant may avoid heat by insulating itself from the heat source; for example, the thick bark insulates the cambium from fire. Some plants tolerate heat by coming to thermal equilibrium with the heat source without suffering injury. Some species evade heat. For example, a plant species that seeds in from unburned areas will have evaded heat stress by escaping the stress.
Numerous workers have identified lethal temperatures for living plant tissue. Hare (1961) and Levitt (1980) published extensive literature reviews that summarized plant response to temperature. Levitt (1980) summarized the lethal high temperature thresholds for different plants and plant parts. Lethal temperatures range from 42 to 70° C for vascular plants, and from 24 to 110° C for nonvascular plants. Conifer seedlings are generally killed by temperatures between 50 and 60° C (Baker 1929; Hare 1961); leaves may be killed at temperatures from 42 to 55° C. Seeds are generally more resistant to heat, with lethal temperatures ranging from 60 to 120° C. Limited studies on heat tolerance of roots (Hare 1961) show that lethal temperatures range from 48 to 54° C.
Factors Influencing Plant Response—A variety of factors influence the lethal temperature of plants and plant parts. Duration of exposure to a given temperature is recognized by most investigators as very important for determining the lethal temperature. Temperature magnitude and duration are controlled by the heat source and the medium of transfer. The lethal temperature of plant tissue varies inversely with exposure time (Hare 1961; Levitt 1980), and the relationship is exponential (Levitt 1980; Wright and Bailey 1982). Baker (1929) observed that heating tissues of conifer seedlings to temperatures of 50° C for long periods did not cause damage, but temperatures a few degrees higher (54° C) caused death within a few minutes. Differential species responses also exist (Baker 1929; Flinn and Pringle 1983).
The metabolic state of the plant and the moisture content of the tissues are also important. Dormant tissues generally tolerate higher temperatures better than actively growing tissues (Hare 1961; Levitt 1980). Some seeds, for example, are able to survive temperatures of 120° C or more in dry conditions and up to 100° C in boiling water, if they have not imbibed water. Lethal temperatures are much lower for seeds with higher moisture contents. In maize seeds, the lethal temperature was 45 to 50° C at moisture contents greater than 40 percent, but was 80° C at moisture contents of less than 10 percent (Levitt 1980). The relationship between moisture and lethal temperature also holds for actively growing tissues (Levitt 1980). These and other data led some investigators to postulate that the moisture content of underground plant parts influences lethal temperature, and further, that dry heat and wet heat may differentially influence lethal temperature (Sapper 1935). The moisture/lethal temperature relationship may be partially explained, because moisture alters the heat-transfer properties and physiological activity of the tissues.
Plant age, vigor, and season have been related to fire response (Noste and Bushey 1987), but it is not clear how responses are related to prefire environmental stress or whether heat resistance is altered. Season of burning influences plant survival and resprouting ability (Flinn and Pringle 1983; Flinn and Wein 1988; Hare 1961; Noste and Bushey 1987). Harrington (1989) postulated that the seasonal effect was related to carbohydrate levels in roots. Fire and other disturbances are most detrimental when carbohydrate reserves in the roots are at the minimum. This is probably directly related to reduction of photosynthetic capacity and the cost of its replacement rather than direct heat damage to roots or sprouting parts. Julander (1945) showed that any treatment favoring accumulation of root reserves increased the growth response of five grass species. Dormant buds are possibly more heat resistant than active buds, thus less susceptible to damage from the downward heat pulse of a fire. Flinn and Pringle (1983) tested the heat tolerance of rhizomes in different seasons, and found that season had a direct effect on the number of sprouts for several species. This response was assumed to be related to stored nutrient reserves.
The size of rhizomes, root crowns, roots, and other structures might influence the amount of heat needed to raise the temperature to lethal levels, thus avoiding the detrimental effects of heat. Flinn and Pringle (1983) studied heat effects on rhizomes, but did not find any relationship between the surface-to-volume ratio and avoidance of high temperatures. Their experimental methods may have masked species differences and effects of surface-to-volume ratio. Larger structures should buffer temperature changes within their tissue. Depending on thermal properties, internal temperatures of these structures may be considerably lower than temperature of the surrounding soil when heat pulses are of short duration.
Species Rooting Habits and Location—Bradley (1984) reviewed a variety of approaches for classifying fire adaptations. Although some of these classifications are not related to fire response the concepts are useful. Raunkiaer's (1934) life-form system and subsequent adaptations (Gill 1981; Keeley 1981; Stickney 1986) accounted for the position of meristematic tissue relative to the ground surface. The life-form system is quite effective for evaluating survival potential, but it does not consider colonization potential, except for residual seed banks within the soil.
In spite of the importance of the morphology and depth of perennating organs, a small number of studies describe the morphology and location of underground regenerating structures. McLean (1969) described structures for plants in forests of British Columbia; Flinn and Wein (1977) described structures in the Acadian Forest of New Brunswick, and several studies described structures for plants in the northwestern United States (Antos and Zobel 1984; Bradley 1984; Miller 1977).
McLean (1969) identified the rooting characteristics of 26 species; Flinn and Wein (1977) sampled the depth of 34 species, and Bradley (1984) evaluated the depth and morphology of eight species. McLean (1969) identified six types of structures based on rooting habit with reproductive tissues located within the duff layers and in the upper 13 cm of mineral soil. Bradley (1984) found rhizomes at depths from 0 to 65 cm in the mineral soil. Flinn and Wein (1977) reported that reproductive structures were found in the litter and duff layers for 50 percent of the species, and below 4 cm for 18 percent of the species. They also found that the depth of reproductive tissue for a particular species varied from 4 to 14 cm between sites and within 2 cm on a site. Both Bradley (1984) and Flinn and Wein (1977) found that depth of reproductive tissues is species specific.
McLean (1969) identified three categories of fire response that are related to rooting structure and depth. Plants with reproductive structures in the litter and duff were susceptible to fire damage. Structures between 1.5 cm and 5 cm deep were intermediate in response and those at depths below 5 cm were not damaged by fire. Bradley (1984) noted sprouting on burned sites from depths as great as 30 cm; she observed that rhizomes with greater mass seemed to have a greater survival potential and deeper rhizomes were more likely to have viable buds after a fire. Flinn and Wein (1977) hypothesized that the deeper the reproductive tissue, the more likely they are to avoid fire damage. They also hypothesized that variations in species response to fire are largely due to interaction between depth (which is species related) and soil heating differences.
Bacteria and Mycorrhizae—The effects of fire on soil microorganisms are complex and varied. Fire impacts microbial numbers and activity directly by heat and in directly by physical and chemical changes in the postfire environment. Generally, the greater the consumption of duff and higher soil heating, the greater the immediate negative impact on the microorganisms. Populations found near the soil surface or within the forest floor were more easily killed than those found in deeper horizons (Ahlgren 1974). Light burning, even on a periodic basis, may have minimal effects on most microbes (Jorgensen and Hodges 1970). Fires of long duration, such as those under slash piles and smoldering duff fires, reduced populations at depths to 25 cm below the surface (Renbuss and others 1973). Actual temperature effects on microbes depend on the type of organism and soil moisture. In general, bacteria are more resistant to heating than fungi. Lethal temperatures for bacteria were reported to be 210° C in dry soils and 110° C in wet soils (Dunn and DeBano 1977). In forest soils, Ahlgren and Ahlgren (1965) found bacteria numbers were reduced by heating to 200° C. Nitrifying bacteria are less resistant than others and were mostly destroyed at 140° C in dry soils and 75° C in moist soils (Dunn and DeBano 1977). Saprophytic fungi in chaparral soils were killed at temperatures above 120° C and 60° C in dry and wet soils, respectively, but "heat shock" fungi prevailed up to 155° C in dry soils and 100° C in moist soils.
Following the initial reduction in microorganism populations, a several-fold increase frequently occurs (Jurgensen and others 1981; Neal and others 1965). Increases are attributed to reinfection of the burned sites and an improved environment for microbial activity, which includes more favorable temperatures, pH, soil moisture, and nutrient levels (Ahlgren 1974). Recovery of nitrifying bacteria appears related to levels of soil heating. Wells and others (1979) reported slow recovery of these bacteria following severe fires, but Jurgensen and others (1981) and White (1986a) found increased populations several weeks after light-to-moderate burns. White (1986b) also indicated that fire may release allelopathic chemicals that inhibit nitrification in ponderosa pine forest floors and soils. Nonsymbiotic nitrogen fixing bacteria also responded positively to burning in several forest types (Lunt 1951).
Soil fungi, which are important in organic matter decomposition and nutrient release, respond similarly to bacteria. An increase in heterotrophic fungi is thought to be responsible for additions of nitrate-N following burning (Wells and others 1979). The response of fungi-producing mycorrhizal roots appears to be related to the prefire nutritional status of the organic and mineral horizons, and to the associated plant species. Mycorrhizal fungi often decrease with burning because these associations occur more frequently in the organic horizons, which can be consumed (Harvey and others 1989). Additionally, mycorrhizae may not form on forest tree seedlings following burning because of increased soil fertility (Wright and Tarrant 1958). However, Black (1985) found more mycorrhizal short roots on Douglas-fir seedlings following a "hard" burn that apparently improved the N status of the soil. Therefore, either increases or decreases in mycorrhizae may be found following burning depending on the degree of soil heating and whether or not critical nutrient levels are achieved.
Root Disease Fungi—Fire is effective in controlling some diseases (Ahlgren 1974; Dickman and Cook 1989; Froehlich and others 1978), but fire may also increase disease by providing infection courts (Ahlgren 1974; Hepting 1935; Littke and Gara 1986; Nordin 1958), by heat stimulation of organisms (Ahlgren 1974), or by providing favorable environments (Ahlgren 1974; Munnecke and others 1981). Most effects of fire are indirect rather than direct. For example, Froehlich and others (1978) were unable to identify the mechanisms by which fire controlled Heterobasidium annosum; but they suspected microclimatic changes, since neither soil pH, chemistry, nor moisture were altered. Thus, the effect appeared to be indirect on the pathogen or the inoculum potential.
The most probable effects of fire on Phellinus weirii are indirect, through the effect on host species dynamics and on the longevity of inoculum. Fire influenced the area and pattern of infestations of P. weirii in a mountain hemlock forest (Dickman and Cook 1989) by reducing the area of infestation, although the pathogen was not eliminated. Fire killed the more susceptible hemlock and led to increased dominance of lodgepole pine, which is fairly resistant to P. weirii damage. Although fire may affect reductions in inoculum by killing the pathogen, enough inoculum may remain to reinvade. Infestation centers may not be manifested everywhere, but the possibility exists that the fungus is hidden (not expressed) in areas where trees are apparently healthy (Dickman and Cook 1989). The lack of expression, or seeming lack of presence, may be due to the fire regime of the area. It is also quite likely that P. weirii-caused mortality creates fuel, enhancing the probability of fire, which, in turn, limits expression of the disease.
Solar Radiation and Soil Temperature—Overstory removal by harvesting or burning increases solar radiation received by the ground surface (Hungerford 1980). Burning not only removes shade, but changes the surface albedo (Hornbeck 1970; Hungerford 1980; McCaughey 1978), which alters the energy balance at the ground surface. Net radiation is greater on burned surfaces than on exposed slash or mineral soil surfaces (Hungerford 1980). Increased solar radiation results in elevated soil temperatures. Exposed soil surface temperatures averaged as much as 36° C warmer at midday and 15° C cooler at night than those on unburned forested sites (Hungerford and Babbitt 1987). These extreme temperatures cause mortality of regenerating tree seedlings (Hare 1961; Shearer 1967). Temperatures in the duff (if it is not consumed) and in the underlying soil are also warmer than those in the undisturbed forest. At a depth of 2 inches, temperatures averaged 7° C warmer (Hungerford and Babbitt 1987). Character of the surface (burned, litter, slash, or bare mineral soil) also significantly influences temperature of the soil to a depth of 16 inches. The duration of these extreme temperature differences is influenced by vegetation recovery rate, which in turn influences regeneration.
Soil Moisture—The moisture status of soils following burning is greatly influenced by organic matter consumption and soil heating. Exposure of burned mineral soil may reduce infiltration and percolation when ash and charred organic matter fill soil pores (Wells and others 1979). Tarrant (1956b) found that percolation rates decreased following severe burns, but increased following light burns. Reduced moisture interception by the forest floor allowed more moisture to reach the mineral soil (Aldon 1968). Haase (1986) reported that soil moisture increased during the growing season following a moderate ponderosa pine underburn. Vegetation removal results in increased soil temperatures and evaporation rates and decreased foliar interception and transpiration losses (Wells and others 1979). The formation of a hydrophobic layer by burning can lead either to a reduction in soil moisture, if the layer is near the soil surface, or to an increase in surface soil moisture if the repellant layer reduces percolation in subsurface horizons.
Plant Regeneration—The reestablishment of vascular plants on burned sites depends on the type, quantity, and morphology of the species present before burning, the characteristics of the fire, and the postfire environment (Morgan and Neuenschwander 1988; Rowe 1983; Stickney 1986). A low-intensity fire that consumes only surface litter fuels generally has little impact on most existing vegetation and results in minimal and short-lived changes in the microenvironment. As fire intensity, fuel consumption, and soil heating increase, resulting in greater damage to vegetation, the contrast in environments from preburn to postburn will also increase.
Natural regeneration of conifers in the northern Rocky Mountains is entirely by seed; therefore, seedbed characteristics are vital to seedling establishment. Seedlings of different conifer species have different environmental requirements for establishment. Generally, seral species establish and grow better than competing climax species in warmer, open environments that result from a fire. Western larch and both seral and climax ponderosa pine regenerate more readily on mineral or lightly duff-covered soils than they do on thick duff (Haase 1986). Thick duff generally dries more rapidly from the surface downward than mineral soil, leaving insufficient moisture for initial seedling root establishment (Boyce 1985). Extreme moisture deficits commonly occur in duff between rains (Potts 1985).
Root productivity is key to survival and growth of plant seedlings. A large root system increases survival potential on drier sites and promotes greater aboveground biomass production on both wet and dry sites by tapping a larger reservoir of nutrients and moisture. Postfire soils with reduced organic horizons influence root activity in several ways. Because root growth is affected by soil temperature, a warmer postburn soil may promote greater root expansion (Hermann 1979). Increased moisture levels in the mineral soil resulting from duff reduction should also favor root growth (Hermann 1979). However, warm and dry conditions on south exposures could be detrimental to root growth and cause root mortality.
Increased soil nutrient concentrations following fire can have various impacts on root response, depending on the initial nutrient status. If nutrients are limiting growth prior to burning, an increase following burning would likely improve root production. If preburn nutrients are less limiting, a large pulse of available nutrients following fire could lead to greater shoot-to-root ratios (Hermann 1979). However, Harrington (1977) found ponderosa pine seedlings growing on burned seedbeds had significantly larger shoots and roots than those on unburned seedbeds. A positive relationship between duff consumption, nutrient levels (especially ammonium-N), and seedling size was implied. Eramian and Neuenschwander (1989) reported greater leader growth of Douglas-fir seedlings on burned sites than on bulldozer-prepared sites, due to warmer soils and more available moisture on the burned sites. Graham and others (1989) emphasized that the degree of burn represented by the amount of duff consumed was important in the evaluation of site preparation for prescribed burning for artificial regeneration. In warm, moist habitat types, burning the surface litter and fine slash fuels, but not the duff layer, was deemed important to optimize growth of planted seedlings.
Reduction of competition is also important for conifer seedling survival and growth. Fire intensity will determine, to a great extent, the type and amount of postfire shrubby and herbaceous competition. Sites with greater soil heating usually are dominated by off site colonizers; Sprouters are common following more moderate fires (Fischer and Bradley 1987; Ryan and Noste 1985). Sprouters are generally more competitive because they are already established. Walstead and Seidel (1990) reported that Douglas-fir seedlings in broadcast-burn clearcuts were larger and had better survival than those in unburned clearcuts. These differences were attributed to reduced competition for the first 3 years of seedling establishment.
Presettlement fire was an important cycling process in forests of the Interior West. Soil properties and plant succession were kept in a dynamic state through fuel consumption and soil heating. Recent forest management activities, primarily fire suppression and harvesting seral species, have led to major changes in many forest types. Organic matter accumulation is greater than in presettlement times, and dense stagnant stands of climax species are common. Because fuel consumption and environmental factors determine the degree of soil heating, which in turn governs the extent of the impact on soil properties, managers must be aware of potential fire effects. Fires that create light to moderate soil heating facilitate nutrient cycling, making nutrients more available to plants and microbes, but soil productivity and stability are adversely affected by excessive heating.
Soil heating (depth and duration of high temperatures) varies widely between fires and within a particular fire because of differences in fuel characteristics, weather conditions, and soil properties (particularly moisture content). Many effects on soil physical, nutrient, and biological properties are related to the amount of organic matter consumed and temperature levels. The physical properties of mineral soil can be changed by heat altering the internal soil structure and by changing the amount and form of carbon compounds. Nutrients can be volatilized, transformed into highly available ions, or remain unchanged by burning. These impacts are directly related to fire-caused temperatures. Temperature ranges and durations also influence survival of microorganisms, plant regenerating parts, and roots. Reestablishment of both macro- and microflora and fauna is dependent on the postfire environment, which, again, is partially determined by the fire effects.
Burning almost always results in redistribution or loss of nutrients (especially N), killing of some plants, changes in postfire environment, and alteration in microbial populations. These changes can be minimized by avoiding fires that cause high soil heating. Prescriptions for sites having shallow, nutrient-poor soils need to be carefully designed and carried out so that nutrient loss and erosion potential are minimized (Grier and others 1989; Harvey and others 1989). In some cases, high soil heating may be needed to reduce fuel accumulations, manipulate plant species to meet a vegetation management objective, or effectively manage a disease problem. Therefore, the objectives should be very clear and the prescriptions should be well designed to provide only the level of soil heating required. Recent research information should help in the development of fire prescriptions that distribute the heat load (Brown and others 1985; Frandsen and Ryan 1986), allow us to predict the level of soil heating (Campbell and others 1990; Hungerford 1990), and assist in the prediction of nutrient changes associated with the predicted soil heating level (Harvey and others these proceedings; Jurgensen these proceedings). Prescribed fire for site preparation and fuel reduction should cause no more disturbance than is actually necessary to accomplish the objectives and maintain or enhance long-term site productivity. Much more research is needed to better define measures of productivity, to apply systems-analysis approaches for solving the complex relations between the factors that influence productivity, and to elucidate the effects of management practices on long-term productivity (Grier and others 1989).
Ahlgren, I. F.; Ahlgren, C. E. 1965. Effects of prescribed burning on soil microorganisms in a Minnesota jack pine forest. Ecology. 46: 304-310.
Ahlgren, Isabel F. 1974. The effects of fire on soil organisms. In: Kozlowski, T. T.; Ahlgren, C. E., eds. Fire and ecosystems. New York: Academic Press: 47-72.
Alban, David H. 1977. Influence on soil properties of prescribed burning under mature red pine. Res. Pap. NC-139. St. Paul, MN: U.S. Department of Agriculture, Forest Service, North Central Forest Experiment Station. 8 p.
Albini, F. A. 1975. An attempt (and failure) to correlate duff removal and slash fire heat. Gen. Tech. Rep. INT-24. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 16 p.
Albini, F. A. 1976. Computer-based models of wildland fire behavior: A users' manual. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 68 p.
Aldon, E. F. 1968. Moisture loss and weight of the forest floor under pole-size ponderosa pine stands. Journal of Forestry. 66: 70-71.
Antos, Joseph A.; Zobel, Donald B. 1984. Ecological implications of below ground morphology of nine coniferous forest herbs. Botanical Gazette. 145(4): 508-517.
Arno, S. F. 1980. Forest fire history in the Northern Rockies. Journal of Forestry. 78(8): 460-465.
Arno, S. F. 1988. Fire ecology and its management implications in ponderosa pine forests. In: Baumgartner, David M.; Lotan, James E., eds. Ponderosa pine: the species and its management: symposium proceedings; 1987 September 29-October 1; Spokane, WA. Pullman, WA: Washington State University, Cooperative Extension: 133-139.
Aston, A. R.; Gill, A. M. 1976. Coupled soil moisture, heat and water vapor transfers under simulated fire conditions. Australian Journal of Soil Research. 14: 55-66.
Ayres, H. B. 1901. Lewis and Clark Forest Reserve, Montana. 21st Annual Report, Part 5. Washington, DC: U.S. Department of the Interior, Geological Survey: 27-80.
Baker, F. S. 1929. Effect of excessively high temperatures on coniferous reproduction. Journal of Forestry. 27: 949-975.
Barrett, Stephen W. 1982. Fire's influence on ecosystems of the Clearwater National Forest: Cook Mountain fire history inventory. Orofino, ID: U.S. Department of Agriculture, Forest Service, Clearwater National Forest, Fire Management. 42 p.
Barrett, S. W.; Arno, S. F. 1982. Indian fires as an ecological influence in the Northern Rockies. Journal of Fores try. 80(10): 647-651.
Bissett, J.; Parkinson, D. 1980. Long-term effects of fire on the composition and activity of the soil microflora of a subalpine, coniferous forest. Canadian Journal of Botany. 58: 1704-1721.
Black, C. H. 1985. Effects and interactions of slash burning and mycorrhizal infection on Douglas-fir seedling growth and morphology. In: Molina, Randy, ed. Sixth conference on mycorrhizae: proceedings; 1984 June 25-29; Bend, OR. Corvallis, OR: Forestry Research Laboratory: 276.
Boyce, Robin B. 1985. Conifer germination and seedling establishment on burned and unburned seedbeds. Moscow, ID: University of Idaho. 68 p. Thesis.
Bradley, Anne F. 1984. Rhizome morphology, soil distribution, and the potential fire survival of 8 species. Missoula, MT: University of Montana. 183 p. Thesis.
Bristow, K. L.; Campbell, G. S.; Papendick, R. I.; Elliott, L. F. 1986. Simulation of heat and moisture transfer through a surface residue-soil system. Agricultural and Forest Meteorology. 36: 193-214.
Brown, J. K. 1985. The "unnatural fuel buildup" issue. In: Lotan, J. E.; [and others], eds. Proceedings-symposium and workshop on wilderness fire. Gen. Tech. Rep. INT-182. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 127-131.
Brown, James K.; See, Thomas E. 1981. Downed dead woody fuel and biomass in the northern Rocky Mountains. Gen. Tech. Rep. INT-117. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 48 p.
Brown, James K.; Marsden, Michael A.; Ryan, Kevin C.; Reinhardt, Elizabeth D. 1985. Predicting duff and woody fuel consumed by prescribed fire in the Northern Rocky Mountains. Res. Pap. INT-337. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 23 p.
Byram, G. M. 1959. Combustion of forest fuels. In: Davis, K. P., ed. Forest fire control and use. New York: McGraw-Hill: 61-89.
Campbell, G. S. 1977. An introduction to environmental biophysics. New York: Springer-Verlag. 159 p.
Campbell, G. S.; Jungbauer, J. D., Jr.; Bidlake, W. R. 1990. Linked transport of heat and water in soil under high temperature (fire) conditions. Unpublished report on file at: U.S. Department of Agriculture, Forest Service, Intermountain Research Station, Intermountain Fire Sciences Laboratory, Missoula, MT. 160 p.
Campbell, R. E.; Baker, M. B., Jr.; Ffolliott, P. F.; Larson, F. R.; Avery, C. C. 1977. Wildfire effects on a ponderosa pine ecosystem: an Arizona case study. Res. Pap. RM-191. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 12 p.
Cattelino, Peter J.; Noble, Ian R.; Slatyer, Ralph O.; Kessell, Stephen R. 1979. Predicting the multiple pathways of plant succession. Environmental Management. 3(1): 41-50.
Chandler, C.; Cheney, P.; Thomas, P.; Trabaud, L.; Williams, D. 1983. Fire in forestry: Volume I Forest fire behavior and effects. New York: Wiley. 450 p.
Chinanzvavana, S.; Grosshandler, W. L.; Davis, D. C. 1986. Analysis of heat transfer in soil during open field burning. Transactions of the American Society of Agricultural Engineers. 29(6): 1797-1801.
Christiansen, E.; Waring, R. H.; Berryman, A. A. 1987. Resistance of conifers to bark beetle attack: searching for general relationships. Forest Ecology and Management. 22: 89-106.
Crookston, N. L.; Stark, R. W. 1985. Forest-bark beetle interactions: stand dynamics and prognoses. In: Waters, W. E.; Stark, R. W.; Wood, D. L., eds. Integrated pest management in pine-bark beetle ecosystems. Wiley-Interscience Publication. New York: John Wiley and Sons: 81-103.
Covington, W. Wallace; Sackett, Stephen S. 1984. The effect of a prescribed burn in southwestern ponderosa pine on organic matter and nutrients in woody debris and forest floor. Forest Science. 30: 183-192.
Covington, W. W.; Sackett, S. S. 1986. Effect of periodic burning on soil nitrogen concentrations in ponderosa pine. Soil Science Society of America Journal. 50: 452-457.
DeByle, Norbert V. 1976. Soil fertility as affected by broadcast burning following clearcutting in northern Rocky Mountain larch/Douglas-fir forests. In: Tall Timbers fire ecology conference No. 14; 1974 October 8-10; Missoula, MT. Tallahassee, FL: Tall Timbers Research Station: 447-464.
DeByle, Norbert V. 1981. Clearcutting and fire in the larch/Douglas-fir forests of western Montana multifaceted research summary. Gen. Tech. Rep. INT-99. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 73 p.
Derby, R. W.; Gates, D. M. 1966. The temperature of tree trunks calculated and observed. American Journal of Botany. 53(6): 580-587.
DeBano, L. F.; Dunn, P. H.; Conrad, C. E. 1977. Fire's effect on physical and chemical properties of chaparral soils. Gen. Tech. Rep. WO-3. Washington, DC: U.S. Department of Agriculture, Forest Service.
DeBano, L. F.; Rice, R. M.; Conrad, C. E. 1979. Soil heating in chaparral fires: effects on soil properties, plant nutrients, erosion, and runoff. Res. Pap. PSW-145. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station. 21 p.
DeVries, D. A. 1958. Simultaneous transfer of heat and moisture in porous media. Transactions, American Geophysical Union. 39(5): 909-916.
Dickman, Alan; Cook, Stanton. 1989. Fire and fungus in a mountain hemlock forest. Canadian Journal of Botany. 67: 2005-2016.
Dyrness, C. T.; Youngberg, C. T. 1957. The effect of logging and slash burning on soil structure. Soil Science Society of America Proceedings. 21: 444-447.
Dunn, Paul H.; DeBano, Leonard F. 1977. Fire's effect on biological and chemical properties of chaparral soils. In: Proceedings of symposium on the environmental consequences of fire and fuel management in Mediterranean ecosystems. Gen. Tech. Rep. WO-3. Washington, DC: U.S. Department of Agriculture, Forest Service: 75-84.
Eramian, Aram; Neuenschwander, Leon F. 1989. A comparison of broadcast burn vs. dozer site preparation methods on the growth of bareroot Douglas-fir seedlings. In: Baumgartner, David M.; Neuenschwander, Leon F.; Wakimoto, Ronald H., eds. Prescribed fire in the Intermountain region: symposium proceedings. Pullman, WA: Washington State University: 75-82.
Federer, C. A. 1982. Subjectivity in the separation of organic horizons of the forest floor. Soil Science Society of America Journal. 46: 1090-1093.
Feller, M. C. 1988. Relationships between fuel properties and slashburning-induced nutrient losses. Forest Science. 34: 998-1015.
Fischer, William C. 1989. The fire effects information system: a comprehensive vegetation knowledge base. In: Ferguson, D. E.; Morgan, P.; Johnson, F. D., compilers. Proceedings land classifications based on vegetation: applications for resource management; 1987 November 17-19; Moscow, ID. Gen. Tech. Rep. INT-257. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 107-113.
Fischer, William C.; Bradley, Anne F. 1987. Fire ecology of western Montana forest habitat types. Gen. Tech. Rep. INT-223. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 95 p.
Flinn, M. A.; Pringle, J. K. 1983. Heat tolerance of rhizomes of several understory species. Canadian Journal of Botany. 61: 452-457.
Flinn, M. A.; Wein, R. W. 1977. Depth of underground plant organs and theoretical survival during fire. Canadian Journal of Botany. 55: 2550-2554.
Flinn, Marguerite A.; Wein, Ross W. 1988. Regrowth of forest understory species following seasonal burning. Canadian Journal of Botany. 66: 150-155.
Frandsen, W. H.; Ryan, K. C. 1986. Soil moisture reduces belowground heat flux and soil temperatures under a burning fuel pile. Canadian Journal of Forest Research. 16: 244-248.
Frandsen, William H. 1989. Smoldering duff: limits, heat evolved, and burn rate. Missoula, MT: University of Montana. 146 p. Thesis.
Frandsen, William H. 1990. Unpublished data on file at: U.S. Department of Agriculture, Forest Service, Inter mountain Research Station, Intermountain Fire Sciences Laboratory, Missoula, MT.
Froelich, R. C.; Hodges, C. S., Jr.; Sackett, S. S. 1978. Prescribed burning reduces severity of Annosus root rot in the South. Forest Science. 24(1): 93-100.
Gates, D. M. 1980. Biophysical ecology. New York: Springer-Verlag. 611 p.
Gill, A. M. 1981. Adaptive responses of Australian vascular plant species to fire. In: Gill, A. M.; Groves, H.; Noble, I. R., eds. Fire and the Australian biota. Canberra, Australia: Australian Academy of Science. 582 p.
Gosz, J. R. 1981. Nitrogen cycling in coniferous ecosystems. In: Clark, F. E.; Rosswall, T., eds. Terrestrial nitrogen cycles. Stockholm: Ecological Bulletin. 33: 405-426.
Graham, Russell T.; Harvey, Alan E.; Jurgensen, Martin F. 1989. Site preparation strategies for artificial regeneration: can prescribed fire fill the bill? In: Baumgartner, David M.; Neuenschwander, Leon F.; Wakimoto, Ronald H., eds. Proceedings, prescribed fire in the Intermountain region: symposium proceedings. Pullman, WA: Washington State University: 83-89.
Grier, C. C. 1975. Wildfire effects on nutrient distribution and leaching in coniferous ecosystems. Canadian Journal of Forest Research. 5: 599-607.
Grier, C. G.; Lee, K. M.; Nadkarni, N. M.; Klock, G. O.; Edgerton, P. J. 1989. Productivity of forests of the United States and its relation to soil and site factors and management practices: a review. Gen. Tech. Rep. PNW-222. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 51 p.
Grime, J. P. 1979. Plant strategies and vegetation processes. New York: John Wiley and Sons. 222 p.
Gruell, G. E. 1983. Fire and vegetative trends in the Northern Rockies: interpretations from 1871-1982 photographs. Gen. Tech. Rep. INT-158. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 207 p.
Gruell, G. E. 1985. Indian fires in the interior West: a widespread influence. In: Lotan, J. E.; Kilgore, B. M.; Fischer, W. C.; Mutch, R. W., tech. coords. Proceedings symposium and workshop on wilderness fire; 1983 November 15-18; Missoula, MT. Gen. Tech. Rep. INT-182. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 68-74.
Gruell, G. E. 1986. Post-1900 mule deer irruptions in the Intermountain West: principle causes and influences. Gen. Tech. Rep. INT-206. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 37 p.
Gruell, G. E.; Schmidt, W. C.; Arno, S. F.; Reich, W. J. 1982. Seventy years of vegetation change in a managed ponderosa pine forest in western Montana implications for resource management. Gen. Tech. Rep. INT-130. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 42 p.
Gruell, G. E.; Brown, J. K.; Bushey, C. L. 1986. Prescribed fire opportunities in grasslands invaded by Douglas-fir. Gen. Tech. Rep. INT-198. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 19 p.
Haase, Sally M. 1986. Effect of prescribed burning on soil moisture and germination on southwestern ponderosa pine seed on basaltic soils. Res. Note RM-462. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 6 p.
Hare, R. C. 1961. Heat effects on living plants. Gen. Tech. Rep. SO-183. New Orleans, LA: U.S. Department of Agriculture, Forest Service, Southern Forest Experiment Station. 32 p.
Harrington, Michael G. 1977. The influence of some environmental factors on initial establishment and growth of ponderosa pine seedlings. Missoula, MT: University of Montana. 107 p. Thesis.
Harrington, Michael G. 1987. Predicting reduction of natural fuels by prescribed burning under ponderosa pine in southeastern Arizona. Res. Note RM-472. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 4 p.
Harrington, Michael G. 1989. Gambel oak root carbohydrate response to spring, summer, and fall prescribed burning. Journal of Range Management. 42(6): 504-507.
Harvey, Alan E.; Jurgensen, Martin F.; Larsen, Michael J.; Graham, Russell T. 1987. Decaying organic materials and soil quality in the Inland Northwest: a management opportunity. Gen. Tech. Rep. INT-225. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 15 p.
Harvey, Alan E.; Jurgensen, Martin F.; Graham, R. T. 1989. Fire-soil interactions governing site productivity in the northern Rocky Mountains. In: Baumgartner, David M.; Neuenschwander, Leon F.; Wakimoto, Ronald H., eds. Prescribed fire in the Intermountain region: symposium proceedings. Pullman, WA: Washington State University: 9-18.
Hepting, G. H. 1935. Decay following fire in young Mississippi Delta hardwoods. Agric. Handb. 386. Washington, DC: U.S. Department of Agriculture, Forest Service. 658 p.
Hermann, R. K. 1979. Growth and production of tree roots. In: Marshall, J. K., ed. The belowground ecosystem: a synthesis of plant-associated processes. Range Science Department Science Serial No. 26. Fort Collins, CO: Colorado State University: 7-28.
Herrington, L. P. 1969. On temperature and heat flow in tree stems. Bull. 73. New Haven, CT: Yale University, School of Forestry. 79 p.
Hornbeck, J. W. 1970. The radiant energy budget of clearcut and forested sites in West Virginia. Forest Science. 16: 139-145.
Hungerford, R. D. 1980. Micro environmental response to harvesting and residue management. In: Environmental consequences of timber harvesting in Rocky Mountain coniferous forests: symposium proceedings; 1979 September 11-13; Missoula, MT. Gen. Tech. Rep. INT-90. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 37-73.
Hungerford, R. D. 1987. Predicting response of understory vegetation to stand treatment: consequences for multi resource management. In: Barger, R. L., compiler. Management of small-stem stands of lodgepole pine workshop proceedings; June 30-July 2, 1986, Fairmont Hot Springs, MT. Gen. Tech. Rep. INT-237. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 118-136.
Hungerford, R. D. 1990. Modeling the downward heat pulse from fire in soils and plant tissue. In: MacIver, D. C.; Auld, H.; Whitewood, R., eds. Proceedings of the 10th conference on fire and forest meteorology; 1989 April 12, 16; Ottawa, ON. Ottawa, ON: 148-154.
Hungerford, R. D.; Babbitt, Ronald E. 1987. Overstory removal and residue treatments affect soil surface, air, and soil temperature: implications for seedling survival. Res. Pap. INT-377. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 19 p.
Jorgensen, J. R.; Hodges, C. S., Jr. 1970. Microbial characteristics of a forest soil after twenty years of prescribed burning. Mycologia. 62: 721-726.
Julander, O. 1945. Drought resistance in range and pasture grasses. Plant Physiology. 20: 573-599.
Jurgensen, Martin F.; Harvey, Alan E.; Larsen, Michael J. 1981. Effects of prescribed burning on soil nitrogen levels in a cutover Douglas-fir/western larch forest. Res. Pap. INT-275. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 6 p.
Keane, R. E. 1987. Forest succession in western Montana computer model designed for resource managers. Res. Note INT-376. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 8 p.
Keeley, J. E. 1981. Reproductive cycles and fire regimes. In: Mooney, H. A.; Bonnicksen, N.; Christensen, J.; Lotan, J.; Reiners, W., tech. coords. Fire regimes and ecosystem properties; proceedings. Gen. Tech. Rep. WO-26. Washington, DC: U.S. Department of Agriculture, Forest Service: 231-277.
Kovacic, D. A.; Swift, D. M.; Ellis, J. E.; Hakonson, T. E. 1986. Immediate effects of prescribed burning on mineral soil nitrogen in ponderosa pine of New Mexico. Soil Science. 141: 71-76.
Leiberg, J. B. 1899. Present condition of the forested areas in Northern Idaho outside the limits of the Priest River Forest Reserve and north of the Clearwater River. 19th annual report. Part V, forest reserves. Washington, DC: U.S. Department of the Interior, Geological Survey: 373-386.
Levitt, J. 1980. Responses of plants to environmental stress. Vol. I chilling, freezing, and high temperature stresses. 2nd ed. New York: Academic Press. 497 p.
Littke, W. R.; Gara, R. I. 1986. Decay of fire-damaged lodgepole pine in south central Oregon. Forest Ecology and Management. 17: 279-287.
Little, Susan N.; Ohmann, Janet L. 1988. Estimating nitrogen lost from forest floor during prescribed fires in Douglas-fir/western hemlock clearcuts. Forest Science. 34: 152-164.
Losensky, B. J. 1990. A comparison of the 1988 fire season to the historical role of fire for the Bob Marshall-Scapegoat Wilderness Complex. Unpublished report on file at: U.S. Department of Agriculture, Forest Service, Lolo National Forest, Missoula, MT.
Lunt, H. A. 1951. Liming and twenty years of litter raking and burning under red and white pine. Soil Science Society of America Proceedings. 15: 381-390.
Lyon, L. Jack. 1966. Initial vegetal development following prescribed burning of Douglas-fir in south-central Idaho. Res. Pap. INT-29. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 17 p.
Lyon, L. Jack; Stickney, Peter F. 1976. Early vegetal succession following large northern Rocky Mountain wildfires. In: Proceedings: Tall Timbers fire ecology conference and Intermountain fire and land management symposium; 1974 October 8-10; Missoula, MT. No. 14. Tallahassee, FL: Tall Timbers Research Station: 355-373.
Martin, R. E. 1963. Thermal properties of bark. Forest Products Journal. 13(10): 419-426.
Martin, R. E. 1982. Fire history and its role in succession. In: Forest succession and stand development research in the Northwest; symposium proceedings. Corvallis, OR: Oregon State University: 92-99.
McCaughey, J. H. 1978. Estimation of net radiation for a coniferous forest, and the effects of logging on net radiation and reflection coefficient. Canadian Journal of Forest Research. 8: 450-455.
McLean, Alastair. 1969. Fire resistance of forest species as influenced by root systems. Journal of Range Management. 22: 120-122.
Miller, Melanie. 1977. Response of blue huckleberry to prescribed fires in a western Montana larch-fir forest. Res. Note INT-188. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 33 p.
Morgan, Penelope; Neuenschwander, Leon F. 1988. Shrub response to high and low severity burns following clearcutting in northern Idaho. Western Journal of Applied Forestry. 3: 5-9.
Munnecke, D. E.; Kolbezen, M. J.; Wilbur, W. D.; Ohr, H. D. 1981. Interactions involved in controlling Armillaria mellea. Plant Disease. 65: 384-389.
Neal, John L.; Wright, Ernest; Bollen, Walter B. 1965. Burning Douglas-fir slash: physical, chemical, and microbial effects in the soil. Forest Lab. Res. Pap. 1. Corvallis, OR: Oregon State University. 32 p.
Niehoff, Gerald J. 1985. Effects of clearcutting and varying fire severity of prescribed burning on levels of organic matter and the mineralization of ammonium nitrogen in the surface layer of forest soils. Moscow, ID: University of Idaho. 43 p. Thesis.
Nissley, Steven D.; Zasoski, Robert J.; Martin, Robert E. 1980. Nutrient changes after prescribed surface burning of Oregon ponderosa pine stands. In: Martin, Robert E.; [and others], eds. Sixth conference on fire and forest meteorology: Proceedings; 1980 April 22-24; Seattle, WA. Washington, DC: Society of American Foresters: 214-219.
Noble, I. R.; Slatyer, R. O. 1977. Post-fire succession of plants in Mediterranean ecosystems. In: Mooney, Harold A.; Conrad, C. Eugene, tech. coords. Proceedings symposium on the environmental consequences of fire and fuel management in Mediterranean ecosystems; 1977 August 1-5; Palo Alto, CA. Gen. Tech. Rep. WO-3. Washington, DC: U.S. Department of Agriculture, Forest Service: 27-36.
Nordin, V. J. 1958. Basal fire scars and the occurrence of decay in lodgepole pine. Forestry Chronicle. 34: 257-265.
Norum, Rodney A. 1976. Fire intensity-fuel reduction relationships associated with understory burning in larch/Douglas-fir. In: Tall Timbers fire ecology conference No. 14; symposium proceedings; 1974 October 8-10; Missoula, MT. Tallahassee, FL: Tall Timbers Research Station: 559-572.
Noste, N. V.; Bushey, C. L. 1987. Fire response of shrubs of dry forest habitat types in Montana and Idaho. Gen. Tech. Rep. INT-239. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 22 p.
Olson, J. S. 1981. Carbon balance in relation to fire regimes. In: Mooney, H. A.; [and others], eds. Fire regimes and ecosystem properties. Gen. Tech. Rep. WO-26. Washington, DC: U.S. Department of Agriculture, Forest Service: 327-377.
Ottmar, R. D.; Little, S. N.; Ohmann, J. 1985. Predicting duff reduction to reduce smoke from clearcut slash burns in western Washington and western Oregon. In: Proceedings of the 8th conference on fire and forest meteorology; 1985 April 29-May 2; Detroit, MI. Washington, DC: Society of American Foresters: 139-144.
Packer, Paul E.; Williams, Bryan D. 1976. Logging and prescribed burning effects on the hydrologic and soil stability behavior of larch/Douglas-fir forests in the northern Rocky Mountains. In: Tall Timbers fire ecology conference No. 14; symposium proceedings; 1974 October 8-10; Missoula, MT. Tallahassee, FL: Tall Timbers Research Station: 465-479.
Packham, D. R. 1969. Heat transfer above a small ground fire. Australian Forest Research. 5: 19-24.
Pafford, D. J. 1986. An investigation into ground surface heating during a prescribed burn. Los Angeles: University of California. 105 p. Thesis.
Pafford, D.; Dhir, V. J.; Anderson, E. B.; Cohen, J. 1985. A model for ground surface heating during a prescribed burn. In: Weather the drive train connecting the solar engine to forest ecosystems: Proceedings of the eighth conference on fire and forest meteorology; 1985 May 2-April 29; Detroit, MI. Washington, DC: Society of American Foresters: 24-32.
Pfister, R. D.; Kovalchik, B. L.; Arno, S. F.; Presby, R. C. 1977. Forest habitat types of Montana. Gen. Tech. Rep. INT-34. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 174 p.
Potts, Donald F. 1984. Water potential of forest duff and its possible relationship to regeneration success in the northern Rocky Mountains. Canadian Journal of Forest Research. 15: 464-488.
Pyne, Stephen J. 1982. Fire in America: a cultural history of wildland and rural fire. Princeton, NJ: Princeton University Press. 654 p.
Raison, R. J.; Woods, P. V.; Jakobsen, B. F.; Bary, G. A. V. 1986. Soil temperatures during and following low-intensity prescribed burning in a Eucalyptus pauciflora forest. Australian Journal of Soil Research. 24(1): 33-47.
Raschke, K. 1960. Heat transfer between the plant and the environment. Annual Review of Plant Physiology. 11: 111-126.
Raunkiaer, C. 1934. The life forms of plants and statistical plant geography. Oxford: Clarendon Press. 632 p.
Renbuss, M.; Chilvers, G. A.; Pryer, L. D. 1973. Microbiology of an ash bed. Proceedings of Linnaean Society of New South Wales: 97: 302-310.
Rowe, J. S. 1983. Concepts of fire effects on plant individuals and species. In: Wein, R. W.; MacLean, D. A., eds. SCOPE 18: The role of fire in northern circumpolar ecosystems. Chichester, NY: John Wiley and Sons: 135-154.
Ryan, K. C. [In press]. Vegetation and wildfire: implications for global climate change. Environment International. Ryan, K. C.; Frandsen, W. H. [In press]. Effects of ground fire on mature ponderosa pine. International Journal of Wildland Fire.Ryan, K. C.; Noste, N. V. 1985. Evaluating prescribed fires. In: Lotan, J. E.; Kilgore, B. M.; Fischer, W. C.; Mutch,R. W., tech. coords. Proceedingssymposium and workshop on wilderness fire; 1983 November 15-18; Missoula, MT. Gen. Tech. Rep. INT-182. Ogden, UT: U.S. Department of Agriculture, Forest Service, Inter mountain Forest and Range Experiment Station: 230-238.
Ryan, K. C.; Reinhardt, E. D. 1988. Predicting postfire mortality of seven conifers. Canadian Journal of Forest Research. 18: 1291-1297.
Sandberg, D. V. 1980. Duff reduction by prescribed burning in Douglas-fir. Res. Pap. PNW-272. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 18 p.
Sandberg, D. V.; Ottmar, R. D. 1983. Slash burning and fuel consumption in the Douglas-fir subregion. In: Seventh conference on fire and forest meteorology symposium: proceedings; 1983 April 25-28; Fort Collins, CO. Boston, MA: American Meteorological Society: 90-93.
Sapper, I. 1935. Versuch zur Hitzresistenz der Pflanzen. Planta. 23: 518-556.
Scotter, D. R. 1970. Soil temperatures under grass fires. Australian Journal of Soil Research. 8: 273-279.
Shearer, R. C. 1967. Insolation limits initial establishment of western larch seedlings. Res. Note INT-64. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 8 p.
Shearer, Raymond C. 1975. Seedbed characteristics in western larch forests after prescribed burning. Res. Pap. INT-67. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 26 p.
Stark, Nellie M. 1977. Fire and nutrient cycling in a Douglas-fir/larch forest. Ecology. 58: 16-30.
Steele, R.; Arno, S. F.; Geier-Hayes, K. 1986. Wildfire patterns change in central Idaho's ponderosa pine-Douglas-fir forest. Western Journal of Applied Forestry. 1(1): 16-18.
Steward, F. R. 1989. Heat penetration in soils beneath a spreading fire. Unpublished paper on file at: U.S. Department of Agriculture, Forest Service, Intermountain Research Station, Fire Sciences Laboratory, Missoula, MT. 6 p.
Stickney, Peter F. 1986. First decade plant succession following the Sundance forest fire, northern Idaho. Gen. Tech. Rep. INT-197. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 26 p.
Tarrant, R. F. 1956a. Effects of slash burning on some soils of the Douglas-fir region. Soil Science Society of America Proceedings. 20: 408-411.
Tarrant, R. F. 1956b. Effects of slash burning on some physical soil properties. Forest Science. 2(1): 18-22.
Van Wagner, C. E. 1972. Duff consumption by fire in eastern pine stands. Canadian Journal of Forest Research. 2(1): 34-39.
Viro, P. J. 1974. Effects of forest fire on soil. In: Kozlowski, T. T.; Ahlgren, C. E., eds. Fire and ecosystems. New York: Academic Press: 7-46.
Walstad, John D.; Seidel, Kenneth W. 1990. Use and benefits of prescribed fire in reforestation. In: Walstad, John D.; Radosevich, Steven R.; Sandberg, David V., eds. Natural and prescribed fire in Pacific Northwest forests. Corvallis, OR: Oregon State University Press: 67-79.
Wells, Carol G.; Campbell, Ralph E.; DeBano, Leonard F.; [and others]. 1979. Effects of fire on soil: a state-of-knowledge review. Gen. Tech. Rep. WO-7. Washington, DC: U.S. Department of Agriculture, Forest Service. 34 p.
Westcot, D. W.; Wierenga, P. J. 1974. Transfer of heat by conduction and vapor movement in a closed soil system. Soil Science Society of America Proceedings. 38: 9-14.
White, C. S. 1986a. Effects of fire on rates of decomposition and nitrogen mineralization in a ponderosa pine ecosystem. Biology and Fertility of Soils. 2: 87-95.
White, C. S. 1986b. Volatile and water-soluble inhibitors of nitrogen mineralization and nitrification in a ponderosa pine ecosystem. Biology and Fertility of Soils. 2: 97-104.
White, E. M.; Thompson, W. W.; Gartner, F. R. 1973. Heat effects on nutrient release from soils under ponderosa pine. Journal of Range Management. 26: 22-24.
Wierenga, P. J.; DeWitt, C. T. 1970. Simulation of heat transfer in soils. Soil Science Society of America Proceedings. 34: 845-848.
Woodmansee, R. G.; Wallach, L. S. 1981. Effects of fire regimes on biogeochemical cycles. In: Mooney, H. A.; [and others], eds. Fire regimes and ecosystem properties. Gen. Tech. Rep. WO-26. Washington, DC: U.S. Department of Agriculture, Forest Service: 379-400.
Wright, E.; Tarrant, R. F. 1958. Occurrence of mycorrhizae after logging slash burning in the Douglas-fir forest region. Res. Note 160. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 8 p.
Wright, Henry A.; Bailey, Arthur W. 1982. Fire ecology: United States and Southern Canada. New York: John Wiley and Sons. 501 p.
Speakers answered questions from the audience after their presentations. Following are the questions and answers on this topic:
Q. Does fire and fire recurrence interval influence the severity of root disease in conifers?
A. A number of associations between fire and root disease can be found in the literature, but causal mechanisms are poorly understood. Fire, or its exclusion, can significantly affect the expression of root disease through its impact on stand structure and species composition. Recurring fire favors seral conifer species that are less susceptible to infection, damage, and mortality from root disease pathogens. The presence of seral species in a healthy stand structure may retard pathogen development and mask root disease expression. Root diseases are generally more prevalent in stands where fire has been excluded for decades. The physical and chemical micro environment is also altered by fire, which can affect population dynamics through innoculum potential and interaction with competitive organisms. Heat generated by fire can be sufficient to kill pathogens in soils and unburned organic matter, but it is not clear what the long-term effects on disease are.
Q. Are there management activities that have effects similar to hot fires? How fast or slow is recovery?
A. No. Management activities other than fire, such as mechanical removal or chemical treatment, may have similar apparent results, but there are considerable qualitative differences. The same amounts of organic material may be treated or removed, which may similarly reduce fire hazard, but great differences in soil chemistry, physical properties, and productivity may occur. No activity other than fire heats the fuel and soil and alters the physical and chemical properties in the same manner. The rate of recovery will depend on the type and intensity of management activity, and is dependent on the specific part of the soil system one is interested in. For example, population levels of some microorganisms may recover and even exceed pretreatment levels within a few weeks of treatment, but nutrients may take years to return to pretreatment levels. More productive sites tend to recover faster than cold, dry, and harsh sites.
Q. Are site preparation/slash burns different than the periodic wildfires historically? Do these two types of fires differ a great deal?
A. Prescribed fire effects can be similar or different than those of historical wildfires. Differences between the two fire types are primarily related to differences in fuel consumption and soil heating, which are influenced by fuel, weather, and soil conditions. On warm dry habitats, historically frequent surface fires maintained light fuel loads. These fires caused minimal soil heating. Under current management, fuel loads on warm, dry habitats are greater after harvesting than commonly occurred historically. Therefore, even though prescribed burns are conducted during more moderate conditions, more fuel is generally consumed, which causes greater heat impacts than occurred during presettlement fires. In more moist habitats, large amounts of fuel accumulated between infrequent historic wildfires. These fires likely caused considerable soil heating. Under current management, fuel loads on moist habitats are similar to presettlement times, but prescribed burns are not conducted during the extremely dry conditions when historic wildfires occurred. Therefore, soil heating and impacts on soils for prescribed fires are likely to be less than for historic wildfires because of differences in fuel, weather, and soil conditions.
Paper presented at the Symposium on Management and Productivity of Western-Montane Forest Soils, Boise, ID, April 10-12, 1990.
Roger D. Hungerford, Michael G. Harrington, and Kevin C. Ryan are research foresters, and William H. Frandsen is a physicist, Intermountain Fire Sciences Laboratory, Intermountain Research Station, Forest Service, U.S. Department of Agriculture, Missoula, MT. Gerald J. Niehoff is a soil scientist, Idaho Panhandle National Forests, Forest Service, U.S. Department of Agriculture, Coeur d'Alene, ID.