- Information by Map Unit
- Information by Topic
- Documents in SoLo
Links to other related sites
- About SoLo
-
- Mission
- Disclaimer
- Authors (complete list)
Understory aids in the restoration of physical, chemical, and biological soil parameters after forest disturbance. Understory establishment must be timely to reduce erosion and maintain forest processes; understory must occur in significant amounts over time to ameliorate disturbance effects, and it must meet management objectives. The goal of vegetative restoration of forest soils is to restore productivity potential to predisturbance levels in a time frame consistent with the cycle of disturbance.
Understory (tree seedlings, shrubs, grasses, and forbs) contributes to the restoration of soil productivity after fire or tree harvest disturbance in western-montane forests. Understory vegetation establishment must be timely to reduce erosion and maintain soil processes and yet not exclude establishment of desirable tree species.
Vegetative restoration encompasses plant structures and processes that maintain or replenish important chemical, physical, and biological soil parameters impacted by disturbance. Early successional species are an integral part of the ecosystem recovery process toward a more stable steady-state condition following disturbance (Marks and Bormann 1972). Natural plant response may be adequate on some sites, but highly disturbed areas may require direct seedings or transplants to maintain soil productivity.
Soil factors frequently affected by disturbance include organic matter content, nutrient content, bulk density and porosity, soil microflora and fauna, and microclimate. Forest disturbance also impacts nutrient cycling, erosion, and soil-development processes.
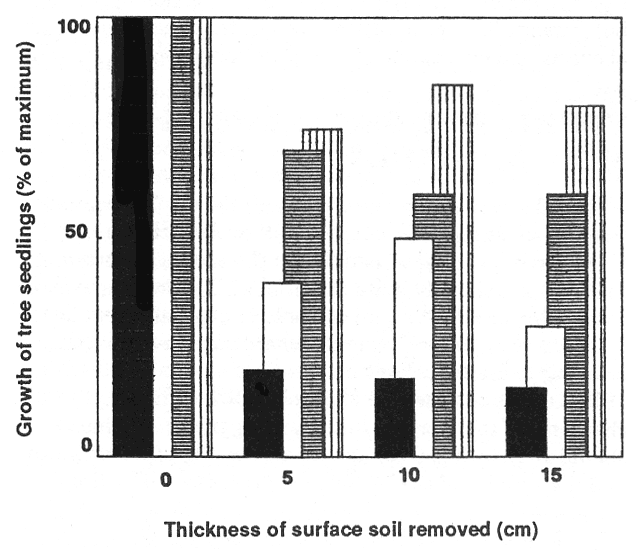
Increased erosion occurs after many types of site disturbance including fire, timber harvesting, residue treatments, and road building (Helvey and others 1985). Soil erosion may occur immediately after removal of the vegetative cover or several years later as roots from the prior vegetation decompose and no longer stabilize the soil (O'Laughlin 1974; Swanston 1974). Loss of the forest floor and surface mineral soil can be a serious obstacle to reforestation or rehabilitation of a site. Removal of the upper soil profile removes most soil organic matter, a large portion of soil nutrients, and beneficial soil biota such as mycorrhizae that are key components to soil productivity (Harvey and others 1980). Bioassay tests indicate significant declines in Douglas-fir (Pseudotsuga menziesii var. glauca [Beissn.] Franco) seedling growth and site productivity with the loss of surface soils, but results differ among soil types (fig. 1, Klock 1982).
Compaction of forest soils often is a problem after tractor harvesting and machine piling of the slash. Compaction causes loss of soil structure, porosity, aeration, and water infiltration. Reduced infiltration rates lead to increased overland flow and accelerated soil erosion. Compaction also limits root growth by reducing pore size, increasing mechanical resistance to root growth, and decreasing oxygen supply. Froehlich (1979) found up to 12 percent reduction in ponderosa pine (Pinus ponderosa Dougl. ex Laws.) growth rates after compaction.
Figure 1—Growth of Douglas-fir seedlings in four eastern Washington soils after removal of successive surface soil layers (Klock 1982). [view larger image - 60K] [Text description of this figure]
Burning results in losses of organic matter, which include both soil organic matter and the aboveground forest biomass with its replenishing inputs of litter for developing soil organic matter. The loss of soil organic matter has detrimental physical, chemical, and biotic effects on soil productivity.
Fire not only eliminates much of the aboveground vegetation but also may eliminate large amounts of the forest floor (Grier 1975). Gessel and others (1973) report the forest floor of the east side of the Cascade Range contains 16,600 to 18,450 kilograms per hectare of organic matter and about 500 to 800 kilograms of nitrogen per hectare. In severe to extreme clearcut and burn treatments, Niehoff (1985) found a 22 to 83 percent reduction in organic matter and an 82 to 99 percent change in mineralizable nitrogen. Leaching losses can further reduce nutrient pools after disturbances, but these losses seem to be relatively small in this region (Clayton and Kennedy 1985; Hart and others 1980).
Many studies have shown that nutrient losses are a frequent result of various site disturbances in montane forests (Clayton 1981; Grier 1975). Removal of the vegetation reduces site nutrients directly through loss of the elements contained in the plant material. For timber harvesting, this can be a substantial amount of some elements (Clayton and Kennedy 1985). Disturbances from logging also can reduce nutrient supplies by erosion losses.
Soil microflora and microfauna populations are controlled by six environmental factors: moisture, temperature, aeration, pH, food substrates, and biological interactions (Harvey and others 1980). Changing any of these factors also can influence the other factors and cause changes in microbial populations (Bollen 1974). Fellin (1980) found clearcutting reduces soil mesofauna populations after harvest in western larch (Larix occidentalis Nutt.)/Douglas-fir stands. Fire generally reduces soil microorganism populations, but many populations increase after temporary reductions (Harvey and others 1989). Fire also can impact ectomycorrhizal fungi infection of root tips, but impacts from clearcutting may differ depending on the aboveground environment (Pilz and Perry 1984).
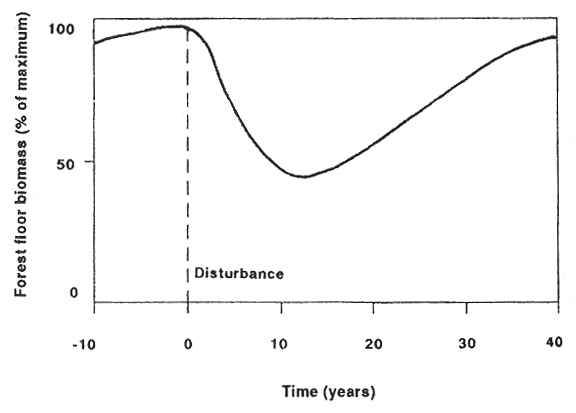
By definition, soil restoration occurs in the "reorganization phase" of stand development after disturbance (fig. 2). Here, forest processes fluctuate as the ecosystem attempts to reestablish steady-state conditions. Disturbance creates a declining cycle in forest processes or structure that is followed by an ascending cycle where parameters are increasing to pre-disturbance levels (Bormann and Likens 1979; Grier and others 1989; Klock 1982). The length and character of the reorganization and aggradation phases should vary with soil parameters, intensity of disturbance, and the activity of the restorative processes (including understory).
For forest floor biomass, the ascending phase may not occur until well after canopy closure occurs (Bormann and Likens 1979); erosion may lessen more rapidly as vegetation occupies the site (Helvey 1980). Vegetation can enhance soil nutrient status and soil processes by organic matter and nutrient inputs from throughfall, litterfall, root exudates, root turnover, N-fixation, and plant senescence and decay.
Vegetation rehabilitation also includes restoring or maintaining favorable soil structure, soil microbial processes, and a favorable microclimate. Although we discuss understory effects on soil restoration in this paper, we recognize that their inputs complement those of green tree retention, forest floor, and woody debris (Wollum and Davey 1978).
Figure 2—The hypothetical reorganization and aggradation phases for the forest floor biomass following tree harvest (adapted from Bormann and Likens 1979). [view larger image - 12K] [Text description of this figure]
Timeliness of vegetation establishment is important in reducing erosion and leaching and in maintaining desirable soil processes (Perry and others 1989). Plants are effective agents for reducing erosion from raindrop impact, overland water flow, and wind (Waldron and Dakessian 1982). Rapid establishment of vegetation enhances capture of nutrients made available by disturbance and prevents their leaching off site (Klock and Grier 1979).
Perry and others (1989) present a good case for reducing intensity of disturbance to maintain understory species that preserve mutualistic soil-plant processes (N-fixation and mycorrhizae-nutrient cycling) from one forest rotation to the next, especially on harsh sites. These "bootstrapping" processes are theoretically maintained across rotations by understory-overstory "species guilds" that share common soil microbial associations. The reestablishment of these mutualistic processes can be difficult on harsh sites if guild species and associated symbionts are not rapidly established. What constitutes a guild member has not yet been defined, but the concept is a "nurse plant" for stand processes.
Duration of understory vegetation in the sere defines the "window" in which understory will make significant contributions to soil productivity. Shrub and herbaceous understory species reach maximum biomass and productivity in early succession and then decline as tree dominance occurs. Snowbrush (Ceanothus velutinus Dougl.) can achieve site dominance within a decade in northeastern Oregon and have an aboveground biomass of 6,560 kilograms per hectare (McNabb and others 1979). Youngberg and Wollum (1976) report snowbrush produced 48,000-54,000 kilograms per hectare of aboveground biomass in a 10-year period. The importance of organic matter inputs from understory exceeds their relative biomass values, in that presence of the lesser vegetation with its litter can enhance microbial activity by faster decomposition and nutrient cycling rates (Youngberg 1965).
Many N-fixing understory species are the first to invade disturbed sites (Klemmedson 1979). Jurgensen and others (1980) identified five genera and seven species on nonleguminous N-fixing plants and eight genera with 25 species of leguminous plants present in the forests of Montana and central Idaho. Pioneer species from the genera of Alnus, Lupinus, Ceanothus, Shepherdia, and Purshia can contribute significant amounts of N during early stand development in specific habitat types, but are absent or under represented on many others (Jurgensen and others 1980).
The amount of N fixed depends on plant density, plant vigor, percentage of plants nodulated, and environmental conditions. Estimates of N-fixation for sitka alder (Alnus sinuata Reg. Rydb.) vary from 17 kilograms per hectare per year to 150 kilograms per hectare per year as shrub density increases from 1,830 to 5,380 and shrub age increases from 3 to 5 years (Heilman and Ekuan 1982). Another nonleguminous N-fixing species, snowbrush, is estimated to provide 72-108 kilograms of nitrogen per hectare per year in ponderosa pine and Douglas-fir stands, respectively (Youngberg and Wollum 1976). Low annual N-fixation rates for bitterbrush (0.57 kilograms of nitrogen per hectare per year) have been reported (Dalton and Zobel 1977). Although amounts of N fixed and released to the soil by snowbrush are under debate (Sylvester 1977), at least a portion of the N in litter (157–178 kilograms of nitrogen per hectare per year) and biomass (295–358 kilograms of nitrogen per hectare per year) (Youngberg and Wollum 1976) will become available to the developing stand at some point. Soils adjacent to snowbrush plants are N-enriched (50-75 percent greater total N) compared to soils where snowbrush does not occur (Binkley and Husted 1983).
Jurgensen and others (1980) suggest snowbrush is the N-fixing species that offers the greatest potential for management. The proposed strategy is to use snowbrush to aid in conifer establishment and for soil N-enrichment followed by spraying to reduce snowbrush competition. Herbicide use is currently restricted, however, so that other alternatives to reduce competition are required. Hand-pulling of 1- to 2-year-old snowbrush seedlings is currently being practiced in the Wenatchee National Forest and is considered economically feasible at a cost of $119–180 per acre, according to Paul Schielke, Forest Silviculturist. Opportunities may exist to achieve a desired density that balances N-enrichment of the site with acceptable competition levels.
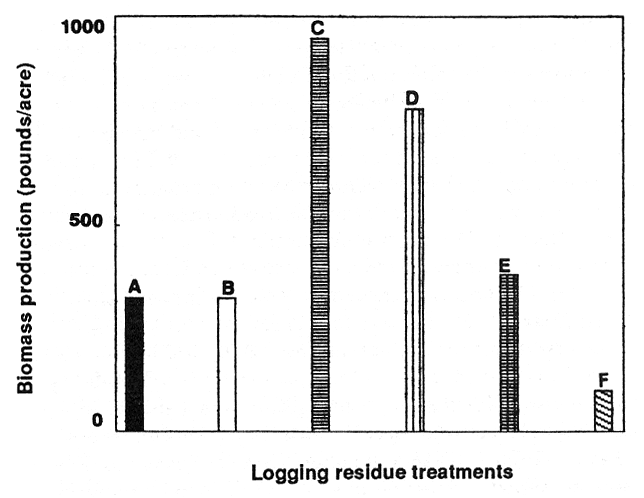
We can manage for increased understory production and potential organic matter accumulation by silvicultural and residue treatments. Schmidt (1980) evaluated the effects of silviculture treatments on understory biomass in a larch and Douglas-fir forest. He found shrub biomass declined from pretreatment values in all treatments, but approached preharvest conditions as intensity of tree harvest increased. He found cover of herbaceous species and small shrubs increased over pretreatment values and were greatest in clearcut treatments. Understory biomass increased over controls when whole-tree harvest was practiced but declined when residues were chipped or burned in lodgepole pine (Pinus contorta Dougl. ex Loud) forests (Packer and Williams 1980; fig. 3). Understory response is species-specific for applied silvicultural treatments (Schmidt 1980).
Future vegetative restoration of soils may become difficult under proposed management strategies such as extended rotations and uneven-aged management that unduly prolong forest stands. Because many of our pioneer species fix N, we need to determine if the longer duration between their normal inputs will impact soil nutrient status. Wollum and others (1968) report the percentage of nodulated snowbrush plants declines exponentially with increasing time between snowbrush crops. Conard and others (1985) suggest this is a result of the absence of the endophyte caused by extended conifer domination of the site.
Figure 3—Understory biomass (pounds per acre) 6 years after tree harvest and residue management treatments in a lodgepole pine forest (Packer and Williams 1980). Residue treatments are (A) control; (B) chipped; (C) residue removed; (D) broadcast burned; (E) piled and burned in rows; and (F) piled and burned between rows. [view larger image - 28K] [Text description of this figure]
Artificial vegetation establishment is warranted when natural response is inadequate to protect the soil resource or when native species are not as desirable. Both direct seedings and transplants have been used for vegetative restoration of disturbed forest soils. Seeding of grass species is recommended for rapid stabilization of disturbed soils, especially by stream channels (Tiedemann and Klock 1976). Seeded grass cover on burned pine and fir forest soils significantly reduced the amount of bare ground in 1 year, but cover of natives was still inadequate by the fourth year on unseeded sites (Anderson and Brooks 1975).
Transplants are another way to rapidly establish plant cover on disturbed sites. Tiedemann and others (1976) were successful in establishing native shrubs on road cut and fill slopes in eastern Washington forests. Established bitterbrush (Purshia tridentata [Pursh] D.C.) and snowbrush may enhance natural succession because of their N-fixing capability. The reader is referred to seeding guides by Hafenrichter and others (1968) and McLean and Bawtree (1971) for species listings and their adaptability to forested sites. Although revegetation is a site-specific process, some general species considerations can be discussed. Grass species most often are used to control erosion on disturbed sites because of ease of establishment and rapid occupation of the site. Klock and others (1975) tested species adaptation of 40 grass, forb, and shrub species on severely disturbed fire lines in east-side Cascade forests. The success of most species decreased with increasing elevation (4,100 to 6,560 feet); this was attributed to freezing, frost heaving, short growing seasons, and high solar radiation levels. Poor performance of seeded grasses has been attributed to low soil N. Legumes have been included in the seed mix (Dyrness 1967) and starter N-S fertilizer has been applied with grass seed on east-side Cascade burn areas (Klock 1969).
Native and introduced legumes have been used in the interior forests as part of erosion control after logging or fire (Jurgensen and others 1980). Legume seedings have been more successful in spring than in fall (Anderson and Elliott 1957), but seeding failures are common (Anderson and Brooks 1975; Brooke and Holl 1988; Dyrness 1967).
Sites with compacted soils require species with roots capable of growth despite high mechanical resistance and poor aeration. Natural or artificial vegetation having this ability and able to provide organic matter to reduce bulk density and stabilize soil structure will be most successful. Alfalfa (Medicago sativa cv. Sonora) roots grew vigorously in compacted soils compared to grasses and ponderosa pine seedlings (Waldron and Dakessian 1982).
Introduction of species into natural ecosystems is an environmental concern in the maintenance of natural vegetative diversity. Helvey and Fowler (1979) found that seeded species reduce native plant cover by one-half (table 1) and adapted seeded species can persist for a decade or more (Geist 1976). Conversely, planting N-fixing species is suggested as a way to facilitate the invasion of native plants having higher soil nutrient requirements (Cargill and Chapin 1987). Although introduced grasses (orchardgrass [Dactylis glomerata L.]) may dominate early plant succession, the probability of their maintaining dominance declines rapidly with stand development, as on the Sleeping Child Burn in western Montana (Lyon 1976).
Vegetative restoration of soils and subsequent competition with tree seedlings remain controversial. Seeding less competitive grasses at 4-5 pounds per acre, followed by intensive grazing, has allowed conifer seedling establishment, but seedling growth was reduced in shelterwood cuts of eastern Oregon (Seidel and others 1990). The long-term benefits of reduced erosion and nutrient inputs must be balanced against reduced tree establishment and growth of tree seedlings that may extend for decades (Schmidt and Shearer 1973). Conversely, seedling establishment can be enhanced by shrub nurse plants (Youngberg 1965), especially on poor sites, in dry years, and with small planting stock (Conard and others 1985).
Both nutrient enrichment and reduced competition effects can be achieved if tree seedlings can be established before understory competition becomes severe (Zavitkovski and others 1969). Mechanisms need to be developed to reduce shrub competition once soil restoration has occurred.
Physical, chemical, and biological attributes of western-montane forest soils can be severely impacted by natural or prescribed fire and practices associated with tree harvesting and residue treatment. Establishment of vegetation is necessary to stabilize the soil surface and initiate processes that replenish nutrient levels and restore or maintain soil physical properties.
Timeliness of vegetation establishment is a factor in reducing erosion and nutrient losses, maintaining soil processes, and maximizing nutrient inputs. Dependency on unpredictable natural vegetation response can be ameliorated by direct seedings or transplants.
Vegetative restoration of soils must consider the maintenance of natural vegetation and safeguard future forest management options. We must also balance the short-term focus of limiting vegetative competition with the broader view of vegetative contributions to long-term site productivity. The defined ecological goal is the restoration or enhancement of soil conditions existing before disturbance in a time frame consistent with the disturbance cycle.
Anderson, C. H.; Elliott, C. R. 1957. Studies on the establishment of cultivated grasses and legumes on burned-over land in northern Canada. Canadian Journal of Plant Science. 37: 97-101.
Anderson, E. William; Brooks, Lee E. 1975. Reduced erosion hazard on a burned forest in Oregon by seeding. Journal of Range Management. 28: 394-398.
Binkley, Don; Husted, Lynn. 1983. Nitrogen accretion, soil fertility, and Douglas-fir nutrition in association with redstem ceanothus. Canadian Journal of Forest Research. 13: 122-125.
Bollen, Walter B. 1974. Soil microbes. In: Cramer, O. P., ed. Environmental effects of forest residues management in the Pacific Northwest: a state-of-knowledge compendium. Gen. Tech. Rep. PNW-24. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station: B1-B41.
Bormann, F. Herbert; Likens, Gene E. 1979. Pattern and process in a forested ecosystem: disturbance, development and the steady state based on the Hubbard Brook ecosystem study. New York: Springer-Verlag. 253 p.
Brooke, R. M.; Holl, F. B. 1988. Establishment of winter versus spring aerial seedings of domestic grasses and legumes on logged sites. Journal of Range Management. 41: 53-57.
Cargill, Susan M.; Chapin, F. Stuart. 1987. Application of successional theory to tundra restoration: a review. Artic and Alpine Research. 19: 366-372.
Clayton, J. L. 1981. Soil disturbances caused by clearcutting and helicopter yarding in the Idaho Batholith. Res. Note INT-305. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 7 p.
Clayton, James L.; Kennedy, Debora A. 1985. Nutrient losses from timber harvest in the Idaho Batholith. Soil Science Society of America Journal. 49: 1041-1049.
Conard, Susan G.; Jaramillo, Annabelle E.; Cromack, Kermit; Rose, Sharon. 1985. The role of the genus Ceanothus in western forest ecosystems. Gen. Tech. Rep. PNW-182. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 72 p.
Dalton, David A.; Zobel, Donald B. 1977. Ecological aspects of nitrogen fixation by Purshia tridentata. Plant and Soil. 48: 57-80.
Dyrness, C. T. 1967. Grass-legume mixtures for roadside soil stabilization. Res. Note PNW-71. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 19 p.
Fellin, David G. 1980. Populations of some forest litter, humus, and soil arthropods as affected by silviculture practices, residue utilization, and prescribed fire. In: Environmental consequences of timber harvesting: symposium proceedings; 1979 September 11-13; Missoula, MT. Gen. Tech. Rep. INT-90. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 287-316.
Froehlich, Henry A. 1979. Soil compaction from logging equipment: effects on growth of young ponderosa pine. Journal of Soil and Water Conservation. 34: 276-278.
Geist, J. Michael. 1976. Forested range fertilization in eastern Oregon and Washington. Rangeman's Journal. 3: 116-118.
Gessel, S. P.; Cole, D. W.; Steinbrenner, E. C. 1973. Nitrogen balances in forest ecosystems of the Pacific Northwest. Soil Biology and Biochemistry. 5: 19-34.
Grier, Charles C. 1975. Wildfire effects on nutrient distribution and leaching in a coniferous ecosystem. Canadian Journal of Forest Research. 5: 599-607.
Grier, Charles C.; Lee, K. M.; Nadkarni, N. M.; [and others]. 1989. Productivity of forests of the United States and its relation to soil and site factors and mangement practices: a review. Gen. Tech. Rep. PNW-222. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 51 p.
Hafenrichter, A. L.; Schwendiman, J. L.; Harris, H. L.; [and others]. 1968. Grasses and legumes for soil conservation in the Pacific Northwest and Great Basin States. Agric. Handb. 339. Washington, DC: U.S. Department of Agriculture, Soil Conservation Service. 69 p.
Hart, George E.; DeByle, Norbert V.; Hennes, Robert W. 1980. Soil solution chemistry and slash disposal. In: Symposium on Watershed Management, Vol 1. New York: American Society of Civil Engineers: 318-325.
Harvey, A. E.; Jurgensen, M. F.; Graham, R. T. 1989. Fire-soil interactions governing site productivity in Northern Rocky Mountains. In: Baumgartner, D.; Breuer, D.; Zamora, B., eds. Prescribed fire in the Intermountain region: Proceedings of the symposium. Pullman, WA: Washington State University: 9-18.
Harvey, A. E.; Jurgensen, M. F.; Larsen, M. J. 1980. Biological implications of increasing harvest intensity on the maintenance and productivity of forest soils. In: Environmental consequences of timber harvesting in Rocky Mountain coniferous forests: symposium proceedings; 1979 September 11-13; Missoula, MT. Gen. Tech. Rep. INT-90. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 211-220.
Helvey, J. D. 1980. Effects of a north central Washington wildfire on runoff and sediment production. Water Resources Bulletin. 16: 627-634.
Helvey, J. D.; Fowler, W. B. 1979. Grass seeding and soil erosion in a steep, logged area in northeastern Oregon. Res. Note PNW-343. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 11 p.
Helvey, J. D.; Tiedemann, A. R.; Anderson, T. D. 1985. Plant nutrient losses by soil erosion and mass movement after wildfire. Journal of Soil and Water Conservation. 40: 168-173.
Heilman, P.; Ekuan, G. 1982. Nodulation and nitrogen fixation by red alder and Sitka alder on coal mine spoils. Canadian Journal of Forest Research. 12: 992-997.
Jurgensen, M. F.; Arno, S. F.; Harvey, A. E.; [and others]. 1980. Symbiotic and nonsymbiotic nitrogen fixation in Northern Rocky Mountain forest ecosystems. In: Gordon, J.; Wheller, C.; Perry, D., eds. Symbiotic nitrogen fixation in temperate forests. Corvallis, OR: Oregon State University Press: 294-308.
Klemmedson, J. O. 1979. Ecological importance of actinoycete-nodulated plants in the western United States. Botanical Gazette. 140: S91-S96.
Klock, G. O. 1969. Use of a starter fertilizer for vegetation establishment. Northwest Science. 43: 38. [Abstract.]
Klock, G. O. 1982. Some soil erosion effects on forest soil productivity. In: Determinants of soil loss tolerance: proceedings of the symposium; 1979 August 5-10; Fort Collins, CO. ASA Spec. Publ. 45. Madison, WI: American Society of Agronomy and Soil Science Society of America: 53-66.
Klock, G. O.; Grier, C. C. 1979. Effects of fire on the long-term maintenance of forest productivity. In: Gessel, S. P.; Kenady, R. M.; Atkinson, W. A., eds. Proceedings, forest fertilization conference; 1979 September 25-27; Union, WA. Contr. 40. Seattle, WA: University of Washington: 247-250.
Klock, G. O.; Tiedemann, A. R.; Lopushinsky, W. 1975. Seeding recommendations for disturbed mountain slopes in north central Washington. Res. Note PNW-244. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 8 p.
Lyon, L. J. 1976. Vegetal development on the Sleeping Child Burn in western Montana, 1961 to 1973. Res. Pap. INT-184. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 23 p.
Marks, P. L.; Bormann, F. N. 1972. Revegetation following forest cutting: mechanisms for return to steady-state nutrient cycling. Science. 176: 914-915.
McLean, A.; Bawtree, A. H. 1971. Seeding forest rangelands in British Columbia. Publ. 1463. Kamloops, BC: Canadian Department of Agriculture. 14 p.
McNabb, D. H.; Geist, J. M.; Youngberg, C. T. 1979. Nitrogen fixation by Ceanothus velutinus. In: Gordon, J.; Wheller, C.; Perry, D., eds. Symbiotic nitrogen fixation in temperate forests. Corvallis, OR: Oregon State University Press: 481-482.
Niehoff, G. J. 1985. Effects of clearcutting and varying severity of prescribed burning on levels of mineralizable nitrogen and organic matter in forest soils. Moscow, ID: University of Idaho. 45 p. Thesis.
O'Laughlin, C. L. 1974. A study of tree root strength deterioration following clearfelling. Canadian Journal of For est Research. 4: 107-113.
Packer, P. E.; Williams, B. D. 1980. Logging residue disposal effects on surface hydrology and soil stability of lodgepole pine forests. In: Environmental consequences of timber harvesting in Rocky Mountain coniferous forests: symposium proceedings; 1979 September 11-13; Missoula, MT. Gen. Tech. Rep. INT-90. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 137-156.
Perry, D. A.; Amaranthus, M. P.; Borchers, J. G.; [and others]. 1989. Bootstrapping in ecosystems. BioScience. 39: 230-237.
Pilz, D. P.; Perry, D. A. 1984. Impact of clearcutting and slash burning on ectomycorrhizal associations of Douglas-fir seedlings. Canadian Journal of Forest Research. 14: 94-100.
Schmidt, W. C. 1980. Understory vegetation response to harvesting and residue management in a larch/fir forest. In: Environmental consequences of timber harvesting in Rocky Mountain coniferous forests: symposium proceedings; 1979 September 11-13; Missoula, MT. Gen. Tech. Rep. INT-90. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 221-248.
Schmidt, W. C.; Shearer, R. C. 1973. Western larch. In: Silviculture systems for the major forest types of the United States. Agric. Handb. 445. Washington, DC: U.S. Department of Agriculture, Forest Service: 56-58.
Seidel, K. W.; Geist, J. M.; Strickler, G. S. 1990. The influence of cattle grazing and grass seeding on coniferous regeneration after shelterwood cutting in eastern Oregon. Res. Pap. PNW-417. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 32 p.
Swanston, D. N. 1974. Slope stability problems associated with timber harvesting in mountainous regions of the Western United States. Gen. Tech. Rep. PNW-21. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 14 p.
Sylvester, W. B. 1977. Dinitrogen fixation by plant association excluding legumes. In: Hardy, R.; Gibson, A., eds. Agronomy and ecology. New York: Wiley: 141-190.
Tiedemann, A. R.; Klock, G. O. 1976. Development of vegetation after fire, seeding and fertilization on the Entiat Experimental Forest. In: Proceedings, Annual Tall Timbers Fire Ecology Conference No. 15; 1974 October; Portland, OR. Tallahassee, FL: Tall Timbers Research Station: 171-192.
Tiedemann, A. R.; Klock, G. O.; Mason, L. L.; Sears, D. E. 1976. Shrub plantings for erosion control in eastern Washington -- progress and research needs. Res. Note PNW-279. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 11 p.
Waldron, L. J.; Dakessian, S. 1982. Effect of grass, legume, and tree roots on soils shearing resistance. Soil Science Society of America Journal. 46: 894-899.
Wollum, A. G.; Davey, C. B. 1978. Nitrogen accumulation, transformation, transport in forest soils. In: Bernier, B.; Winget, C., eds. Forest soils and forest land mangement. Quebec: Laval University Press: 67-106.
Wollum, A. G.; Youngberg, C. T.; Chichester, F. W. 1968. Relation of previous timber stand age to nodulations by Ceanothus velutinus. Forest Science. 14: 114-118.
Youngberg, C. T. 1965. Silviculture benefits from brush. Detroit, MI: Proceedings, Society of American Foresters: 55-59.
Youngberg, C. T.; Wollum, A. G. 1976. Nitrogen accretion in developing Ceanothus velutinus stands. Soil Science Society of America Journal. 40: 109-112.
Zavitkovski, J.; Newton, M.; El-Hassan, B. 1969. Effects of snowbrush on growth of some conifers. Journal of Forestry. 242-247.
Paper presented at the Symposium on Management and Productivity of Western-Montane Forest Soils, Boise, ID, April 10-12, 1990.
Richard Everett and Darlene Zabowski are Plant Ecologist and Soil Scientist, respectively, Forestry Sciences Laboratory, Pacific Northwest Research Station, Forest Service, U.S. Department of Agriculture, 1133 N. Western Ave., Wenatchee, WA 98801. Phillip McColley is Soil Scientist (retired), USDA Forest Service, Wenatchee, WA.