- Information by Map Unit
- Information by Topic
- Documents in SoLo
Links to other related sites
- About SoLo
-
- Mission
- Disclaimer
- Authors (complete list)
This paper is a synthesis of published research pertinent to soils developed from Mount Mazama tephra deposits in the Pacific Northwest. The research discussed includes studies of physical and chemical properties, of changes in these properties imposed by forest harvest, of relations between altered properties and tree growth, and of vegetation responses to fertilization.
Recent and historical volcanic activity has deposited ash and pumice on portions of the western United States and affected the productivity of some forested areas. Our paper focuses primarily on soils developing in ejecta from Mount Mazama in Oregon. We have utilized research findings pertinent to these soils as examples of what may exist in soils developed from other pumice and ash sources, both local and more general. We also recognize that considerable variability is likely in these widely occurring parent materials.
Our objectives are to overview productivity relationships of these soils to physical and chemical properties, to harvest-induced changes in these properties, and to soil fertility management in tests of nutrient additions from fertilizers.
McCool (1914) made early observations of physical and chemical properties of pumice soils in central Oregon. He found these soils had high porosity, high water-holding capacity, and rapid infiltration rates. According to Fryxell (1965) the eruption of Mount Mazama occurred some 6,600 years ago and affected about 900,000 km2 to the north and east of the source, now known as Crater Lake, in Oregon. Particle size and thickness generally decline with increasing distance from the source. Particles near the source are highly porous, gravelly pumice but are dominantly silt sized at the extremities of the deposition. The ejecta near the source formed a pumice-mantled plateau of about 50,000 km2 in central Oregon. At greater distance from the source, the finer deposition also imposed important changes, but the soils are distinctly different from pumice soils (Geist and Strickler 1978).
About 64 km (40 miles) from the source a typical soil profile has a sandy loam A1 horizon about 5 cm thick, a 10- to 30-cm-thick loamy sand AC, a 70-cm gravelly loamy sand C1, and a 50+-cm loamy sand C2 over a IIB and/or IIC buried horizon. This soil and closely associated soils of the pumice-mantled plateau have recently been classified as Xeric Vitricryands.
The C1 horizon contains coarser textured material of special interest, because the individual pumice particles are bridged together resembling a three-dimensional jigsaw puzzle. The bridging restricts root proliferation (Cochran 1971; Hermann and Petersen 1969). Sinker roots of ponderosa pine (Pinus ponderosa Dougl. ex Laws.) usually penetrate through the C1 horizon, often through old root channels and rodent burrows. Roots of lodgepole pine (Pinus contorta Dougl. ex Loud.) are mainly restricted to the A1 and AC horizons. Because limited root penetration of the C1 and C2 horizons severely alters the positional availability of both water and nutrients, productivity is lowered where these horizons are present. The restricted rooting also makes trees more vulnerable to windthrow.
At greater distance from the source the mantle of pumice is thinner and the particle size smaller. The AC horizon becomes thicker, the C1 thins or disappears, and restrictions on root growth lessen or become nonexistent. At these distances more mixing of weathered and unweathered material has apparently resulted from windthrow, animal burrowing, and redistribution by water and wind action. Because of this alteration, the C1 horizon is mixed in varying degrees with finer material, so particle bridging is much reduced, and productivity is generally higher than where mixing has not occurred.
An older, preeruption, buried soil occurs below the Mazama pumice layer at depths varying from 50 cm to over 3 m. Hermann and Petersen (1969) found sharp increases in height growth of young ponderosa pine (8-20 feet tall) where root systems penetrated the C1 and C2 horizons and reached the buried soil profile.
Thermal properties of these soils combined with climatic conditions of the area result in frequent radiation frost during the growing season. Frost heaving can also be a severe problem in limiting natural regeneration and can completely eliminate a fall planting.
Radiation frost damage is a general problem in the pumice zone because of the high elevation, the dry air mass, the predominance of clear night skies, and the thermal properties of the soils. Pumice soils have thermal properties that more closely resemble peat soils than common mineral soils. Thermal conductivities, thermal diffusivities, and thermal contact coefficients are low. Therefore, surface temperatures are extremely warm in the day and quite cold at night (Cochran 1975b). Damage from radiation frost is accentuated in basins and flats where cold air accumulates. Lodgepole pine is better suited to these areas than ponderosa pine because of greater tolerance of germinating lodgepole seedlings to temperatures below 7.8°C (Cochran and Bertnsen 1973), and because the female cones of lodgepole pine are less susceptible to damage at low temperatures (Sorensen and Miles 1974). Ponderosa pine is more predominant on raised areas or uplands (Dyrness and Youngberg 1966; Youngberg and Dyrness 1959), and the scattered ponderosa pine in lodgepole areas on flats and basins apparently becomes established only beneath the shelter of larger lodgepole pine. Young ponderosa pines in basins and flats require sheltering until they reach about 2 m height. Planting of ponder osa pine in these areas without overstory protection results in stunted plants (Cochran 1984). Natural regeneration difficulties will be extreme after clearcutting, and even lodgepole pine may require planting if regeneration is desired within 5 years. Pumice deserts created by cutting or fire are hard to artificially regenerate, and natural regeneration could take many years. An ecologically more compatible silvicultural strategy could be to maintain lodgepole pine and use shelterwoods without complete slash cleanup to help regenerate new stands. Such an approach will perpetuate the existence of an onsite seed source of lodgepole pine for ensuing crops, while providing some protection to scattered ponderosa pine which may occur (Cochran 1975b). Conversions to pure ponderosa pine should be avoided.
Frost heaving is another problem accentuated by the soils and climate of the pumice-mantled plateau. Frost heaving begins when soils are moist to the surface and day-to-night temperatures fluctuate somewhat above and below freezing. Water in the surface layer freezes rapidly after sunset, creating an upward gradient in soil water potential. Water continues to move upward and freeze, and the pore space in a thin soil layer at the surface becomes filled with ice. As more water continues to move up from the unfrozen layer and freezes at the ice-water interface, ice crystals elongate downward and separate the frozen and unfrozen soil layers. Plants with root collars encased in the frozen layer can be completely or partially heaved. Fall planting will be completely heaved if not covered immediately by snow, which persists until spring. Therefore, only spring planting is recommended. To protect natural seedlings from frost heaving some uncrushed logging slash should be retained on the soil surface. Shelterwood cutting offers little, if any, protection from frost heaving problems, but does reduce radiation frost problems (Cochran 1975b).
At 400 km (250 miles) from the source, finer particle deposition produced ash soils that lack the severe thermal problems inherent in central Oregon pumice soils, and forest associations differ as well. In eastern Oregon and Washington, western and northern Idaho, western Montana, and southern British Columbia and Alberta, volcanic ash soils are associated with several forest types including: lodgepole pine, western larch (Larix occidentalis Nutt.), Douglas-fir (Pseudotsuga menziesii var. glauca [Biessn.] Franco), grand fir (Abies grandis [Dougl. ex D. Don] Lindl.), mixed conifer, spruce-fir, and others. Mixed conifer may include grand fir, interior Douglas-fir, western larch, subalpine fir (Abies lasiocarpa [Hook.] Nutt.), western white pine (Pinus monticola Dougl. ex D. Don), and Engelmann spruce (Picea engelmannii Parry ex Engelm.) in eastern Oregon and Washington. Farther east and north, western hemlock (Tsuga heterophylla [Raf.] Sarg.) and western redcedar (Thuja plicata Donn ex D. Don) may also occur in the mixed conifer type. Ash soils are usually associated with the most productive forest sites.
Ash profiles in eastern Oregon commonly have 50-60 cm of light-colored, weakly structured, silt-loam textured ash overlying 20+ cm of a darker colored buried soil of residuum or colluvium commonly from basalt or andesite. The buried material is silty clay loam to clay loam in texture. No severe rooting restrictions are naturally present. These ash soils and related andic intergrades include Vitrandepts, Cryandepts, and Eutroboralfs. A common profile sequence has 5-10 cm of A horizon, 30-40 cm AC, 10-20 cm C, overlying 10-20 cm of IIB and IIC. Colors in the ash overburden lighten markedly with drying. In northern Idaho and northwestern Montana ash overlies granitic materials, including those weathered from intrusive rock or compacted glacial till, the latter being restrictive to rooting. Some of these soils have been reported as Dystrochrepts, Dystrandepts, and Fragiochrepts (Niehoff 1985; Nielson-Gerhardt 1986).
Some of the distinctive properties of volcanic ash and pumice soils are evident when compared to basalt-derived soils (table 1). The 0- to 30-cm depth of ash and pumice soils has lower bulk density, is lower in clay content, has fewer coarse fragments, and has higher potentially available soil-water storage. A high water-holding capacity, roughly twice that of an ordinary coarse-textured soil, is one of the unique features McCool (1914) found in early investigations of pumice soils. The clays of ash and pumice soils are dominantly amorphous, but they do contain minor amounts of layer silicates (Chichester and others 1969; Fosberg and others 1979; Harward and Youngberg 1969).
Ash and pumice, which have low volume weight and relatively few profile coarse fragments, can easily be moved by heavy equipment. Consequently, their vulnerability to mechanical displacement is high, especially when soil moisture levels are low. Even though they have naturally low bulk densities, ash and pumice soils will compact to growth-affecting levels, as is discussed further in subsequent sections. Under natural conditions ash and pumice soils generally have moderate to high infiltration rates and are well drained (McCool 1914).
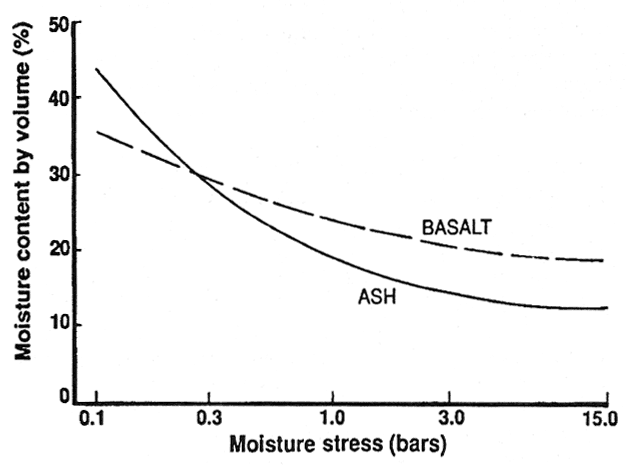
Desorption studies of soil moisture content at various soil moisture tensions show ash materials have higher water content at lower tensions, similar water content at higher tensions, and yield more water in the lower tension range, as compared to basalt-derived materials (fig. 1). Given fully charged profiles, volcanic ash can supply more readily available water to plants than can basalt-derived soils (Geist and Strickler 1978). The relation between moisture content and moisture tension for pumice soils is very similar to that of ash soils, although the relation does vary among pumice horizons because of different amounts of vesicular pumice particles that exceed 2 mm in diameter. These coarser materials add to the water-holding capacity of the profile, but they exhibit slow unsaturated water movement because of disrupted pore continuity between and within pumice particles (Cochran 1966; Youngberg and Dyrness 1964).
Figure 1—Generalized relationships of volumetric moisture content to soil moisture stress in volcanic ash- and basalt-derived soils (adapted from Cochran 1966; Geist and Strickler 1978; Youngberg and Dyrness 1964). [view larger image - 20K] [Text description of this figure]
Chemical properties will be addressed in relation to vegetation to illustrate some associations with chemical attributes. In central Oregon, both ponderosa and lodgepole pines are associated with pumice soils. In eastern Oregon, ponderosa pine is more often associated with basalt- and andesite-derived soils; while lodgepole pine, western larch, interior Douglas-fir, mixed conifer, and spruce-fir types are associated with volcanic ash soils (Geist and Strickler 1978).
As in most soils, organic matter, total N, and extractable P are higher in the surface layers of ash and pumice soils and decline with depth (table 2). This orientation appears to be exaggerated, though in varying degree, in pumice soils. Extractable cations are less surface oriented, and in fact, commonly occur in higher concentrations, deeper in the profile. Youngberg and Dyrness (1964) found that profile quantities (kg/ha) of extractable cations could increase 1.5-2 times where thicker pumice C horizons occurred. Geist and Strickler (1978) found concentrations (meq/100 g) of bases in the buried soil were commonly twice those of the ash material immediately above.
The effects of vegetation, parent material, and climate seem to be evident in these chemical data. Youngberg and Dyrness (1964) found snowbrush (Ceanothus velutinus), a known nitrogen fixer, enriched the surface 2 cm of pumice soils with N. Calcium was also higher in the surface. Geist and Strickler (1978) found vegetation- associated differences in bases among ash soils (L, M, SF in table 2), with those supporting spruce-fir being much lower. In contrast, organic matter and total N levels were much higher. Spruce-fir sites are associated with wetter and cooler conditions, which are conducive to leaching of bases and slowed decomposition of organic matter. Unfortunately, we cannot separate vegetation and climatic influences.
| Vegetation1 | Depth cm |
Organic matter Percent |
Total N Percent |
Extractable P ppm |
Extractable Ca meq/100g |
|---|---|---|---|---|---|
| 1L = lodgepole pine, M = mixed conifer, SF = spruce-fir, P = ponderosa pine, B = bitterbrush (Purshia tridentata [Pursh] DC.), in P/S and W/S the S = snowbrush (Ceanothus velutinus Dougl.), W = white fir or grand fir. | |||||
| Ash-Derived Soils of Eastern Oregon | |||||
| L | 0–15 | 3.60 | 0.09 | 51 | 4.2 |
| L | 15–30 | 1.60 | .05 | 31 | 4.3 |
| M | 0–15 | 3.43 | .10 | 63 | 8.2 |
| M | 15–30 | 1.93 | .06 | 36 | 6.8 |
| SF | 0–15 | 7.89 | .22 | 45 | 3.0 |
| SF | 15–30 | 4.70 | .16 | 21 | 1.8 |
| Pumice-Derived Soils of Central Oregon | |||||
| P/B | 0–2 | 6.41 | 0.14 | 12 | 2.4 |
| P/B | 2–10 | 1.57 | .03 | 6 | 1.2 |
| P/S | 0–2 | 9.54 | .17 | 19 | 4.3 |
| P/S | 2–10 | 1.23 | .04 | 7 | 1.4 |
| W/S | 0–2 | 8.84 | .23 | 20 | 6.1 |
| W/S | 2–10 | 1.17 | .03 | 9 | 2.0 |
| Basalt-Derived Soils of Eastern Oregon | |||||
| P/B | 0–15 | 4.08 | 0.11 | 44 | 10.7 |
| P/B | 15–30 | 2.16 | .07 | 31 | 9.9 |
Basalt-derived soils supporting ponderosa pine in eastern Oregon had similar organic matter, total N, and extractable P levels, but were generally higher in extractable base levels. These soils were geographically associated with the ash soils supporting lodgepole pine and mixed conifer types, so macroclimatic influences should not differentially affect soil chemistry; but we still have confounded effects of vegetation and parent material.
Youngberg and Dyrness (1964) suggested soil chemical data should be compared on a mass per unit area (volume) basis to account for large disparities in bulk density. Comparisons of classical kinds of data such as in table 2 are valid among soils of similar bulk densities. For purposes of broader comparison we include values representative of the upper 60 cm (2 ft) of mineral soil for three macronutrients (table 3). These values further indicate pumice soils are on the low end of extractable bases and P range (as are ash soils under spruce-fir), but have moderate levels of nitrogen (depending on positional availability to root systems). Youngberg and Dyrness also noted the upper 60 cm of mineral soil supporting medium-site Douglas-fir in western Oregon could contain about 8,960 kg/ha N and proportionately higher levels of other nutrients compared to pumice soils. Differences in comparison with ash soils are not always this great for some nutrients, but there are some large differences in nutrient capital (Geist and Strickler 1978).
| Nutrient | Ash-derived | Pumice | Basalt | |||
|---|---|---|---|---|---|---|
| L1 | M | SF | P | P | ||
1L = lodgepole pine, M = mixed conifer, SF = spruce-fir, P = ponderosa pine. |
||||||
| Total N | 1,973 | 2,157 | 5,263 | 1,992 | 2,292 | |
| Extr. P | 110 | 132 | 94 | 31 | 105 | |
| Extr. K | 1,135 | 1,521 | 755 | 614 | 1,458 | |
Our tabular data do not include contributions from organic horizons, which are often thin. However, litter layers are generally known to supply significant levels of nutrients. The contribution of nutrients from litter is probably more important now than historically, since fire suppression and improved grazing practices have aided litter accumulations. Nielson-Gerhardt (1986) found substantial amounts of nitrogen mineralization in ash soils, as is noted later. Roots of ponderosa and lodgepole pine have been observed in litter layers of pumice soils. The presence of these roots may be further indication that nutrient supply from organic layers is important.
Potential nitrogen availability in volcanic ash soils before logging has been measured using laboratory assessments of nitrogen mineralization. Anaerobic test values averaged 9 ppm N for surface mineral layers in northern Idaho under mixed conifers (Niehoff 1985). Under mixed conifers in northwestern Montana, test values ranged from 10 to 16 ppm N in surface mineral layers and 48 to 93 ppm in organic layers (Nielson-Gerhardt 1986). In eastern Oregon and Washington, values for surface mineral layers of ash soils averaged 10 ppm under various conifer cover, about half the levels of basalt-derived soils (Geist unpublished data). Values for surface mineral layers of another group of 14 ash soils associated with several overstory types ranged from 8 to 44 ppm N and averaged 19 ppm (Geist 1977). These latter values were well above the average extractable ammonium N (7 ppm) and nitrate N (2 ppm).
The foregoing discussions of physical and chemical properties indicate volcanic ash and mixed ash-pumice depositions represent a forest-growth-enhancing addition to the prior soil; but where pumice soils lack mixing in the C1 and C2 horizons (nearer the source), a less favorable growth medium exists after deposition.
The potential for adverse effects on forest growth associated with mechanical impacts on soil properties has concerned both foresters and soil scientists for decades. Relatively few studies of mechanical impacts on ash and pumice soils of interior forests have been conducted; but the findings, so far, are similar to those obtained elsewhere on other soils.
Investigations of tractor harvesting on 11 units with ash soils in eastern Oregon and southeastern Washington showed that 10 to 36 percent of the area of logging units sustained excessive compaction. Excessive compaction was defined as a 20 percent increase in bulk density (Geist and others 1989). These data excluded area in the transportation system but included skid trails and landings. Notably, there were some units where significant percentages of area were excessively compacted, but changes in average bulk density were relatively small.
Sullivan (1988) monitored harvest units to assess soil impacts from single and multiple forest management entries. He found eight of 13 units with ash soils had detrimental impacts significantly exceeding 20 percent of the areas (P ≤ 0.10). Nearly all of the detrimental impacts were associated with excessive compaction.
Cochran and Brock (1985) investigated relations between soil compaction and tree growth in a deep, well-drained soil formed from Mount Jefferson ash 50 to 60 cm thick over residuum or colluvium. They found both total height and height growth of trees in 5-year-old ponderosa pine plantations decreased with increasing bulk densities. When contrasting high and low soil disturbance classes in silt loam ash soils of eastern Washington and Oregon, Geist and others (unpublished) found reduced height and radial growth of lodgepole and ponderosa pine associated with compaction and soil displacement.
In the mixed conifer type of northern Idaho, Nielson-Gerhardt (1986) took 0- to 10-cm samples of volcanic ash soils to assess the effects of logging disturbance on estimates of nitrogen availability using anaerobic incubation tests. Test values for samples from severely disturbed sites were 50 percent lower (5-10 ppm N) than samples from relatively undisturbed sites within the same clearcuts. Because the severely disturbed category was missing an organic layer, no mineralized N was available from that source. Severely disturbed areas constituted 25-50 percent of the four clearcuts studied. Organic matter differences were not detected, but a high degree of mixing and variability was associated with disturbances. On the average, ash thickness was cut in half (loss ranged from 10 to 15 cm) because of displacement and compression. The rooting medium for new tree seedlings was sharply reduced, because the underlying glacial till was ice compacted and restrictive to root growth. Severe disturbance categories had 19 to 58 percent higher bulk densities in the ash layer than unlogged forest controls.
Niehoff (1985) found no significant differences in organic matter content or nitrogen mineralization tests of volcanic ash soils except in the extremely burned category 1 year after clearcutting and burning mixed conifer stands in northern Idaho. The extreme category had lost all the organic layer and was pink to red 1 to 5 cm into the mineral surface layer. Less than 1 ppm N was mineralized in the extreme category; other categories, including a forested control, had 9 to 10 ppm of N mineralized from 2.5 to 7.5 cm samples of anaerobically incubated mineral soil.
In studies of ash-influenced soils of north-central Idaho, Clayton and others (1987) measured diameter, radial growth, and height of trees 15-25 years old in relation to soil displacement and compaction assessments. One or more growth variables were significantly reduced in response to changes in displacement or compaction disturbance classes. The trees were naturally regenerated lodgepole pine or planted ponderosa pine.
Compaction was found to persist for decades in volcanic-influenced soils studied by Froehlich and others (1985) and in silt loam ash studied by Geist and others (1989). Froehlich and others found persistence depended mainly on the initial degree of change, which in turn may be influenced in varying degree by soil texture, structure, moisture conditions, number of machine passes, loading, and operator skills. No difference was found in the rate of recovery between two different textured soils.
From the foregoing it is evident that, even though ash and pumice soils have relatively low bulk densities before and after disturbance, there is a strong association between growth loss and compaction. In addition, there is evidence to suggest other disturbance influences, such as displaced soil that removes onsite nutrients and water-holding capacity, may reduce growth. More attention is warranted, not only to compaction, but to these other influences.
Pumice and ash soils in south-central Oregon have generally low fertility levels. Nitrogen (N), phosphorus (P), and sulfur (S) seem to be the limiting nutrients. Our view is that S and P appear to be more limiting to grasses than to trees, but our experience is limited to relatively few members of the ash and pumice soil populations.
Greenhouse studies of nutrient additions to ash soils (0- to 15-cm mineral depths) found strong N and S interaction responses in grass growth. There were indications that N additions without S could reduce growth (Geist 1971). Tests of elemental S versus calcium sulfate, in combination with either ammonium nitrate or urea sources of N, showed the sulfate source was clearly superior for stimulating growth of grass (Geist 1976a; Klock and others 1971).
Pumphrey (1971) annually fertilized a northeastern Oregon ash soil for 4 consecutive years with 67 kg N, 11 kg P, and 12 kg S/ha. Dry-matter production of 28 grass species was increased an average of 2,016 kg/ha. Growth began earlier and green period extended longer with fertilization. Geist (1976b) compared production of newly seeded orchardgrass (Dactylis glomerata L.) on ash soil using various fertilizers at a rate of 100 kg N/ha. First-year dry matter yield for the unfertilized control was 213 kg/ha; the N-only treatment from ammonium nitrate yielded 372 kg/ha, and the N plus S treatment from ammonium sulfate yielded 1,490 kg/ha. Subsequent research on ash soil fertility has shown varying degrees of N, S, and P deficiencies and thus variable nutrient interactions (Geist 1977, 1979).
When pine seedlings are grown in the greenhouse on material from the AC horizon of the Lapine soil (pumiceous, Xeric Vitricryands) in central Oregon, responses to N, P, and S and all the interactions were significant (P ≤ 0.05, Youngberg and Dyrness 1965). Fertilizer ratios of N:P:S that appeared to produce the best seedling response in the greenhouse were 200:100:33. Boron (B) and other micronutrient deficiencies have been suspected, but greenhouse or field trials have not confirmed these suspicions. Thinned stands of ponderosa or lodgepole pine show significant responses (P ≤ 0.05) in bolewood growth when fertilized with N, P, and S in the above ratios at rates of N ranging from 224 to 672 kg/ha (Cochran 1978, 1979, 1989). Lodgepole pine fertilized at the highest rate produced 131 percent in gross periodic annual volume increment (PAI) over the controls during the first 4-year period after application. Ponderosa pine fertilized at the lowest level produced a 56 percent increase in gross PAI over the controls during the first 5 years. Much of this response appears to be related to N, and fertilization with N alone at a rate of 224 kg/ha produces a significant growth response. S appears to be the next most limiting nutrient (Will and Youngberg 1978).
A field study of N, P, and S combinations showed basal area growth of thinned ponderosa pine was superior using N plus P plus S compared to N plus S, N plus P, or N alone (Cochran 1978). Similar superiority was not found with bolewood, perhaps because it is more difficult to measure bolewood as precisely. Thinned white fir (Abies concolor [Gord. & Glend.] Lindl.) on pumice soils fertilized at rates of 224 kg/ha N plus 40 kg/ha S experienced more than a 32 percent increase in gross PAI over a 5-year study period (Cochran 1990). Thinned interior Douglas-fir responds well to fertilization with 224 kg/ha N on ash soils in eastern Washington and northern Idaho, but increasing the rate to 448 kg/ha N did not yield additional response. Similar results were obtained in thinned ponderosa pine on ash soils of eastern Washington and northeastern Oregon. Fertilized western larch in northeastern Washington produced 27 percent more gross PAI than controls during an 8-year period after fertilization (Cochran and Vander Ploeg 1988).
Grasses growing under pine stands on the pumice-mantled plateau also respond to fertilization. Growth of western needlegrass (Stipa occidentalis Thrub. ex Wats.), and bottlebrush squirreltail (Sitanion hystrix [Nutt.] J.G. Smith) under a thinned lodgepole overstory was significantly increased (P ≤ 0.05) by fertilization with 672 kg N plus 336 kg P plus 111 kg S/ha (Cochran 1975a). Lower rates of nutrients in other studies on the pumice-mantled plateau also appeared to increase grass production, but the response was not quantified (Cochran 1978, 1979). Responses of the shrub understory to fertilization of pumice soils were inconsistent, apparently varying by individual plant. Leader growth of some bitterbrush (Purshia tridentata [Pursh] DC.) plants under either lodgepole or ponderosa pine appeared to respond to fertilization, but others did not. Measuring the response of bitterbrush plants in the field was further complicated by selective browsing of big game. Some bitterbrush plants were heavily browsed, while others nearby appeared untouched. Fertilized plots of Idaho fescue (Festuca idahoensis Elm.) in the understory of ponderosa and lodgepole pine were also heavily grazed by game, but control plots were hardly grazed at all. Geist and others (1974) found fertilization increased big game preference for grass growing in eastern Oregon ash soils.
Understanding the beneficial, detrimental, and in some cases, unique properties of volcanic ash and pumice soils is critical to maintaining the long-term productivity of interior forests in tephra deposition zones. Mismanagement of these soils can easily occur. If it does, full rehabilitation may not be possible, and productivity will be reduced.
Chichester, F. W.; Youngberg, C. T.; Haward, M. D. 1969. Clay mineralogy of soils formed on Mazama pumice. Soil Science Society of America Proceedings. 33: 115-120.
Clayton, J. L.; Kellogg, G.; Forrester, N. 1987. Soil disturbance-tree growth relations in central Idaho clearcuts. Res. Note INT-372. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 6 p.
Cochran, Patrick H. 1966. Heat and moisture transfer in a pumice soil. Corvallis, OR: Oregon State University. 165 p. Dissertation.
Cochran, P. H. 1971. Pumice particle bridging and nutrient levels affect lodgepole and ponderosa pine seedling development. Res. Note PNW-150. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 10 p.
Cochran, P. H. 1975a. Response of pole-sized lodgepole pine to fertilization. Res. Note PNW-247. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 10 p.
Cochran, P. H. 1975b. Soil temperature and natural forest regeneration in south-central Oregon. In: Bernier, B.; Winget, C. H., eds. Forest soils and forest land management: Proceedings of the Fourth North American Forest Soils Conference; 1973 August. Quebec City, Quebec, Canada: Laval University Press: 37-52.
Cochran, P. H. 1978. Response of a pole-sized ponderosa pine stand to nitrogen, phosphorus, and sulfur. Res. Note PNW-319. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 8 p.
Cochran, P. H. 1979. Response of thinned ponderosa pine to fertilization. Res. Note PNW-339. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 8 p.
Cochran, P. H. 1984. Should ponderosa pine be planted on lodgepole pine sites? Res. Note PNW-419. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 6 p.
Cochran, P. H. 1989. Growth rates after fertilizing lodgepole pine. Western Journal of Applied Forestry. 4(1): 18-20.
Cochran, P. H. 1990. [In press]. Response of thinned white fir stands to fertilization with nitrogen plus sulfur. Manuscript submitted for publication. Copy on file at the Silviculture Laboratory, 1027 NW Trenton Avenue, Bend, OR 97701.
Cochran, P. H.; Bertnsen, Carl M. 1973. Tolerance of lodgepole and ponderosa pine seedlings to low temperatures. Forest Science. 19(4): 272-280.
Cochran, P. H.; Boersma, L.; Youngberg, C. T. 1967. Thermal properties of a pumice soil. Soil Science Society of America Proceedings. 31(4): 454-459.
Cochran, P. H.; Brock, Terry. 1985. Soil compaction and initial height growth of planted ponderosa pine. Res. Note PNW-434. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 4 p.
Cochran, Pat; Vander Ploeg, Jim. 1988. Some response results for lodgepole pine, grand fir, western larch and ponderosa pine. In: Moore, Jim, compiler. 1988. Unpublished proceedings: Intermountain forest tree nutrition cooperative: a workshop for member organizations; 1988 April 12; Moscow, ID. Copy on file at the Silviculture Laboratory, 1027 NW Trenton Avenue, Bend, OR 97701.
Dyrness, C. T.; Youngberg, C. T. 1966. Soil-vegetation relationships within the ponderosa pine type in the central Oregon pumice region. Ecology. 47: 122-138.
Fosberg, M. A.; Falen, A. L.; Jones, J. P.; Singh, B. B. 1979. Physical, chemical and mineralogical characteristics of soils from volcanic ash in northern Idaho: I. Morphology and genesis. Soil Science Society of America Journal. 43: 541-547.
Froehlich, H. A.; McNabb, D. H. 1984. Minimizing soil compaction in Pacific Northwest forests. In: Stone, Earl L., ed. Forest soils and treatment impacts: Proceedings Sixth North American Forest Soils Conference; 1983 June 19-23; Knoxville, TN. Knoxville, TN: University of Tennessee, Department of Forestry, Wildlife and Fisheries: 159-192.
Froehlich, H. A.; Miles, D. W. R.; Robbins, R. W. 1985. Soil bulk density recovery on compacted skid trails in central Idaho. Soil Science Society of America Journal. 49: 1015-1017.
Fryxell, R. 1965. Mazama and Glacier Peak volcanic ash layers: relative ages. Science. 147: 1288-1290.
Geist, J. M. 1971. Orchardgrass responses to fertilization of seven surface soils from the central Blue Mountains of Oregon. Res. Pap. PNW-122. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 12 p.
Geist, J. Michael. 1976a. Orchardgrass growth on nitrogen and sulfur fertilized volcanic ash soil. Journal of Range Management. 29(5): 415-418.
Geist, J. M. 1976b. Forested range fertilization in eastern Oregon and Washington. Rangeman's Journal. 3: 116-118.
Geist, J. M. 1977. Nitrogen response relationships of some volcanic ash soils. Soil Science Society of America Journal. 41(5): 966-1000.
Geist, J. M. 1979. Toward improved understanding of N and S fertility status of upland volcanic ash soils of eastern Oregon and Washington. Agronomy Abstracts: 171.
Geist, J. M.; Edgerton, P. J.; Strickler, G. S. 1974. "Yukky to yummy" with fertilizers. Rangeman's Journal. 1: 39-41.
Geist, J. M.; Strickler, G. S. 1978. Physical and chemical properties of some Blue Mountain soils in northeast Oregon. Res. Pap. PNW-236. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station. 19 p.
Geist, J. M.; Hazard, J. W.; Seidel, Kenneth W. 1989. Assessing physical conditions of some Pacific Northwest volcanic ash soils after forest harvest. Soil Science Society of America Journal. 53(3): 946-950.
Harward, M. E.; Youngberg, C. T. 1969. Soils from Mazama ash in Oregon: identification, distribution and properties. In: Pedology and quaternary research symposium: Proceedings; 1969 May 13-14. Edmonton, AB: University of Alberta: 163-178.
Hazard, J. W.; Geist, J. M. 1984. Sampling forest soil conditions to assess impacts of management activities. In: Stone, Earl L., ed. Forest soils and treatment impacts: Proceedings, Sixth North American Forest Soils Conference; 1983 June 19-23; Knoxville, TN. Knoxville, TN: University of Tennessee, Department of Forestry, Wildlife and Fisheries: 421-430.
Hermann, Richard K.; Petersen, Roger G. 1969. Root development and height increment of ponderosa pines in pumice soils of central Oregon. Forest Science. 15: 226-237.
Klock, G. O.; Geist, J. M.; Tiedemann, A. R. 1971. Erosion control fertilization from pot study to field testing. Sulphur Institute Journal. 7: 7-10.
McCool, M. M. 1914. Some unusual soils that occur in Oregon. Agronomy Journal. 6: 159-164.
Niehoff, G. J. 1985. Effects of clearcutting and varying severity of prescribed burning on levels of organic matter and the mineralization of ammonium nitrogen in the surface layer of forest soils. Moscow, ID: University of Idaho. 43 p. Thesis.
Nielson-Gerhardt, M. 1986. Effects of logging and mechanical site preparation on levels of ammonium nitrogen and organic matter in forest soils. Moscow, ID: University of Idaho. 74 p. Thesis.
Pumphrey, F. V. 1971. Grass species growth on a volcanic ash derived soil cleared of forest. Journal of Range Management. 24: 200-203.
Sorensen, Frank C.; Miles, Richard S. 1974. Differential frost tolerance of ponderosa and lodgepole pine megasporangiate strobili. Forest Science. 20(4): 377-378.
Sullivan, T. E. 1988. Monitoring soil physical conditions on a national forest in eastern Oregon: a case study. In: Slaughter, Charles W.; Gasbarro, T., eds. Proceedings of the Alaska Forest Soil Productivity Workshop; 1987 April 28-30; Anchorage, AK. Gen. Tech. Rep. PNW-219. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: 69-76.
Will, G. M.; Youngberg, C. T. 1978. Sulfur status of some central Oregon pumice soils. Soil Science Society of America Journal. 42(1): 132-134.
Youngberg, C. T. 1959. The influence of soil conditions, following tractor logging, on the growth of planted Douglas-fir seedlings. Soil Science Society of America Proceedings. 23: 76-78.
Youngberg, C. T. 1975. Effects of fertilization and thinning on the growth of ponderosa pine. Soil Science Society of America Proceedings. 39: 137-139.
Youngberg, C. T.; Dyrness, C. T. 1959. The influence of soils and topography on the occurrence of lodgepole pine in central Oregon. Northwest Science. 33: 111-120.
Youngberg, C. T.; Dyrness, C. T. 1964. Some physical and chemical properties of pumice soils in Oregon. Soil Science. 97: 391-399.
Youngberg, C. T.; Dyrness, C. T. 1965. Biological assay of pumice soil fertility. Soil Science Society of America Proceedings. 29(2): 182-187.
Speakers answered questions from the audience after their presentations. Following are the questions and answers on this topic:
Q. What is the relative compaction hazard of ash cap soils compared to sands, silts, and glacial tills?
A. There appears to be considerable similarity among textural groups in their response to compaction, if one expresses compaction in terms of percentage increase in bulk density. Thus, the absolute values of bulk densities before and after compaction are less important than the percentage change. This view follows that of Froehlich and McNabb (1984), who provided a more detailed explanation with appropriate qualifiers. It has been my feeling that volcanic ash soils compact quite easily (readily compact), but I haven't seen engineering studies that would bear out my viewpoint. Regardless, ash soils do not achieve particularly high bulk densities (are not highly compactable). I suggest you also consult Geist and others (1989).
Q. Does compaction of soils make trees more susceptible to insect and disease?
A. I believe this is the case. However, I have not conducted studies in this regard. There are other papers in this proceedings that may offer you specific answers. My view is that compaction (site stress) increases physiological stress in trees which in turn increases vulnerability to insect and disease.
Paper presented at the Symposium on Management and Productivity of Western-Montane Forest Soils, Boise, ID, April 10-12, 1990.
J. M. Geist and P. H. Cochran are Research Soil Scientists, Forestry and Range Sciences Laboratory, 1401 Gekeler Lane, La Grande, OR 97850, and Silviculture Laboratory, 1027 NW Trenton, Bend, OR 97701, respectively; Pacific Northwest Research Station, Forest Service, U.S. Department of Agriculture.